Article Text

Abstract
Background/aims To identify KIF11 mutations in patients with familial exudative vitreoretinopathy (FEVR) and to describe the associated phenotypes.
Methods Mutation analysis in a cohort of patients in a single institute was conducted. Bioinformatics was performed for whole exome sequencing, and the variants were confirmed by Sanger sequencing. Clinical data and DNA samples were collected from 814 unrelated Chinese probands, including 34 with FEVR, at the Pediatric and Genetic Eye Clinic, Zhongshan Ophthalmic Centre, Guangzhou, China.
Results Four novel heterozygous truncation mutations in KIF11, including c.131_132dupAT (p.P45Ifs*92), c.2230C>T (p.Q744*), c.2863C>T (p.Q955*) and c.2952_2955delGCAG (p.G985Ifs*6), were detected in four of 34 probands with FEVR. Combined with our previously identified mutations in FEVR cases (n=14), KIF11 mutations were identified in 8.3% (4/48) of all probands with FEVR. Ocular phenotypes documented in patients with KIF11 mutations showed a significant great variability of FEVR from the avascular zone in the peripheral retina to bilateral complete retinal detachment. Analysis of available family members in family QT1314 and QT937 showed segregation of KIF11 mutations with the phenotype of FEVR as expected. The family QT964 with two affected siblings and unaffected parents demonstrated a peculiar somatic mosaicism in the mother who had a low copy number variant (about 7% in her leucocyte DNA).
Conclusions Identification of mutations in 8.3% patients suggests KIF11 mutations as a common cause of FEVR. Patients with KIF11 mutations showed typical, but variable, signs of FEVR with or without microcephaly, lymphoedema and mental retardation.
Statistics from Altmetric.com
Introduction
Familial exudative vitreoretinopathy (FEVR, MIM 133780), a rare hereditary disease causing blindness, is characterised by developmental anomalies of the peripheral retinal vasculature.1 Aberrant vascularisation results in various fundus changes, including non-perfusion in the peripheral retina, straightening of the temporal arcade, retinal neovascularisation, vitreoretinal traction, retrolenticular fibrotic mass, retinal fold and tractional retinal detachment.2 Variable clinical manifestations are presented in patients with FEVR, ranging from asymptomatic peripheral vascular anomalies to congenital blindness due to complete retinal detachment.3 FEVR can be inherited as autosomal dominant,1 ,3 autosomal recessive,4 and X-linked trait,5 in which the autosomal dominant form is the most common.
Mutations in six genes have been reported to be responsible for FEVR, including FZD4,6 LRP5,7 TSPAN12,8 NDP,5 ATOH79 and ZNF408.10 Mutations in these genes have been identified in about 50% cases of FEVR,11 suggesting a genetic basis for the rest half cases remains to be identified.
Mutations in KIF11 were reported to be associated with microcephaly with or without chorioretinopathy, lymphoedema or mental retardation (MLCRD, MIM152950), in which, unclassified chorioretinal dysplasia was identified in about half of patients.12 ,13 Recently, KIF11 mutations were first identified in patients with FEVR by Robitaille et al.14 The purpose of this study was to estimate the contribution of KIF11 mutations in our patient cohort of FEVR and the associated phenotypes.
In this study, potential pathogenic variants in KIF11 were selected from whole exome sequencing on 814 Chinese probands with different forms of eye diseases, including 34 with FEVR. Then, the variants were confirmed by Sanger sequencing and further evaluated in the available family members. Four novel heterozygous truncation mutations were identified, and the associated phenotypes were documented.
Methods
Subjects
The study was approved by the institutional review board of the Zhongshan Ophthalmic Centre, and written informed consents were collected from the participants or their guardians before the study. In total, 814 probands with different forms of genetic eye diseases, including 34 with FEVR, were recruited at the Pediatric and Genetic Eye Clinic, Zhongshan Ophthalmic Centre. In the current study, the 34 probands with FEVR included 11 newly recruited and 23 for whom mutations in the four known FEVR genes (FZD4, LRP5, TSPAN12, and NDP) had been excluded by Sanger sequencing in our previous studies.15–17 Totally, mutations in the four known FEVR genes were identified in three probands of the 11 new recruited and 14 in our previous studies.15–17 Of the 34 families, six had a family history of FEVR with an autosomal dominant trait and 28 were isolated cases. FEVR was diagnosed based on the presence of retinal vascular developmental anomaly as previously described.17 Genomic DNA of the probands and the available relatives was extracted from leucocytes of the peripheral blood as previously described.18 Fundus fluorescein angiography was used to validate the diagnosis of FEVR in a suspected case. Following the discovery of KIF11 mutations, head circumference, lymphoedema and mental development were assessed in patients with KIF11 mutations.
Whole exome sequencing
Genomic DNA from the 814 probands was initially analysed by whole exome sequencing, which was performed by Marcogen (http://www.macrogen.com/).19 Briefly, genomic DNA was captured using the Illumina TruSeq Exome Enrichment Kit (62 M) and exome-enriched DNA fragments were sequenced on the Illumina HiSeq2000. The average sequencing depth was 125-fold. The high quality reads were mapped to the human genomic reference sequence hg19 using BWA (http://bio-bwa.sourceforge.net/) and variants were detected through SAMtools (http://samtools.sourceforge.net/).
Variants analysis
Initially, all variants in KIF11 were collected from the data based on whole exome sequencing of 814 probands. Multi-bioinformatics analysis was performed to filter the potential pathogenic variants as follows: (1) excluding variants in non-coding regions and synonymous variants in exonic regions without affecting splicing site predicted by the Berkeley Drosophila Genome Project (http://www.fruitfly.org/seq_tools/splice.html); (2) excluding variants with minor allele frequency >0.01 by Exome Aggregation Consortium (http://exac.broadinstitute.org/gene/ENSG00000138160); (3) excluding variants predicted to be benign by both PolyPhen-2 (http://genetics.bwh.harvard.edu/pph2/) and Sorting Intolerant From Tolerant (http://sift.jcvi.org/www/SIFT_enst_submit.html). The remaining variants were considered as pathogenic candidates.
Sanger sequencing
Candidate pathogenic variants in KIF11 were validated by Sanger sequencing. The targeted fragments containing variants were amplified by a touchdown PCR as previously described.20 The primers were designed using the online tool Primer3 (http://primer3.ut.ee/) (table 1). The amplicons were analysed using an Applied Biosystems (ABI) 3130 Genetic Analyzer (Applied Biosystems, Foster City, California, USA) with BigDye Terminator cycle sequencing kit V.3.1. Variants were identified on the SeqManII program of the Lasergene package (DNAStar, Madison, Wisconsin, USA) by aligning the amplicon sequences with consensus sequences from the National Centre for Biotechnology Information human genome database (http://www.ncbi.nlm.nih.gov/). Validated variants were further analysed in available family members.
Primers used in Sanger sequencing
Genotyping
Genotyping was performed on four available members of family QT964. Three 5′-fluorescently labelled microsatellite markers adjacent to KIF11 were selected from the ABI PRISM Linkage Mapping Set MD-10 (Applied Biosystems). Multiplex PCR was performed as previously described.21 PCR products were mixed with GENESCAN 400 HD (ROX) standards (ABI) and deionised formamide, denatured at 95°C for 5 min. The amplicons were separated on an ABI 3130 Genetic Analyzer (Applied Biosystems) and then analysed using the Gene Mapper software package V.4.0 (Applied Biosystems). Haplotype was generated using the Cyrillic V.2.1 program (Cyrillic Software, Wallingford, UK). The markers' information was obtained from the Genethon database (http://www.bli.uzh.ch/BLI/Projects/genetics/maps/gthon.html).
Cloning sequencing
The target fragments covering the mutation sites of KIF11, c.2230C>T from the leucocyte DNA of the QT964I:2 or c.2952_2955delGCAG from QT761I:1 and QT761I:2, were amplified using PCR with the primers KIF11-E17 and KIF11-E21, respectively. The fragments were cloned into pMD19-T simple vectors (Takara BIO, Japan) according to the manufacturer's instructions. The resultant plasmids were transformed into Escherichia coli JM109 for amplification. The plasmids were isolated from the suspension. Fragments covering the mutant allele were amplified by PCR and Sanger sequencing was used to confirm the mutant and wild type alleles. The positive recombinants containing the mutant allele as well as those with the normal allele were counted, respectively. The mutant allele frequency was calculated as the ratio of the number of the recombinants containing the mutant allele to that of the total positive ones.
Results
Mutations detected
Four novel heterozygous truncation variants in KIF11 were detected through whole exome sequencing in four of 34 probands with FEVR: c.131_132dupAT (p.P45Ifs*92), c.2230C>T (p.Q744*), c.2863C>T (p.Q955*) and c.2952_2955delGCAG (p.G985Ifs*6) (table 2 and figure 1). The detection rate of KIF11 mutations in patients with FEVR was 8.3% (4/48) and 12.9% (4/31) of the probands without the known FEVR gene mutations. These four variants were then confirmed by Sanger sequencing. None of these four mutations were presented in the other 780 inhouse controls. Of the four mutations, two were nonsense and two were frameshift mutations. Of the four families with KIF11 mutations, three families had a familial history of FEVR, and one was an isolated case (figure 1). Segregation analysis in family QT1314 and QT937 suggested that mutations in KIF11 segregated with the phenotypes of FEVR.
KIF11 mutations identified in the families with FEVR

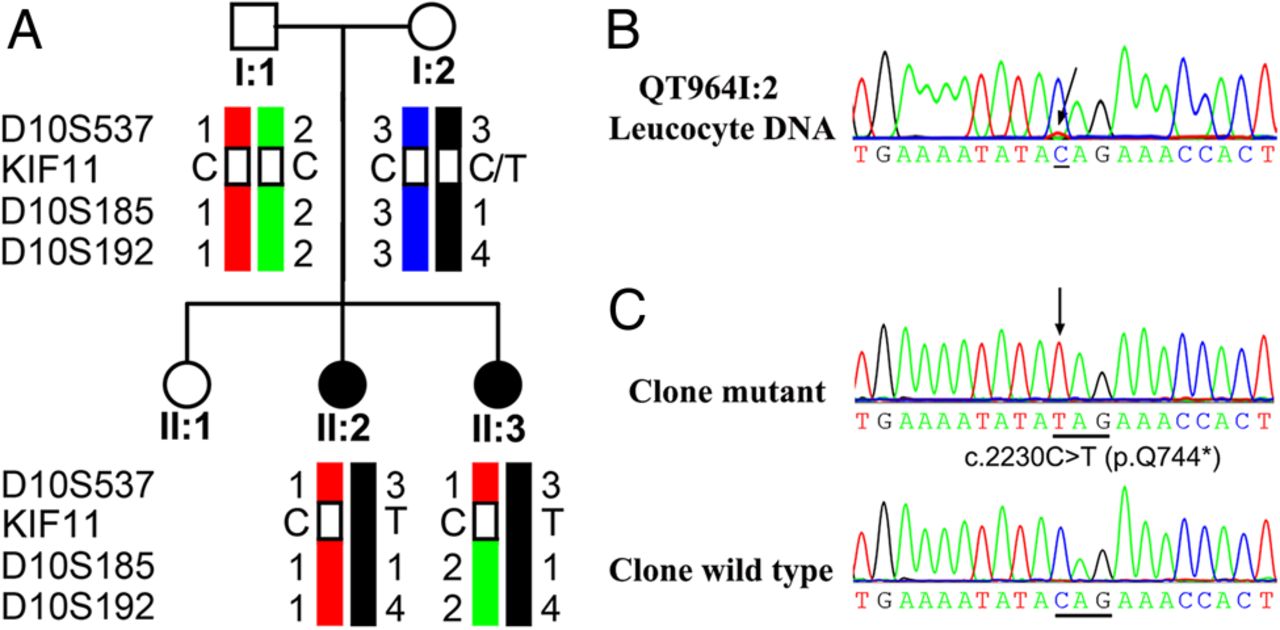
Pedigrees and Sequence chromatography. The columns from left to right display the family number, pedigree, sequence chromatography from patients and controls. In the pedigrees, M sign represents a variant; +, a normal allele; arrows, probands; squares, males; circles, females; blackened symbols, affected individuals. In the sequence chromatography, the variations are marked with arrows and named under the sequence.
Two missense variants, c.994A>G (p.I332V) and c.2747A>G (p.D916G), were detected, but considered to be less likely causes of FEVR (see online supplementary table S1) for the following reasons: (1) the two missense variants presented in four probands with different ocular diseases: p.I332V in one with FEVR and one with cone dystrophy, p.D916G in one with congenital stationary night blindness in whom Nyctalopin c.121delG had been identified as causative,22 and one with glaucoma; (2) segregation analysis showed that the two variants in KIF11 were present in unaffected as well as affected individuals (see online supplementary figure S1). Clinical information of the variant carriers was present in the online supplementary table S2 and figure S2.
Phenotypic characteristics
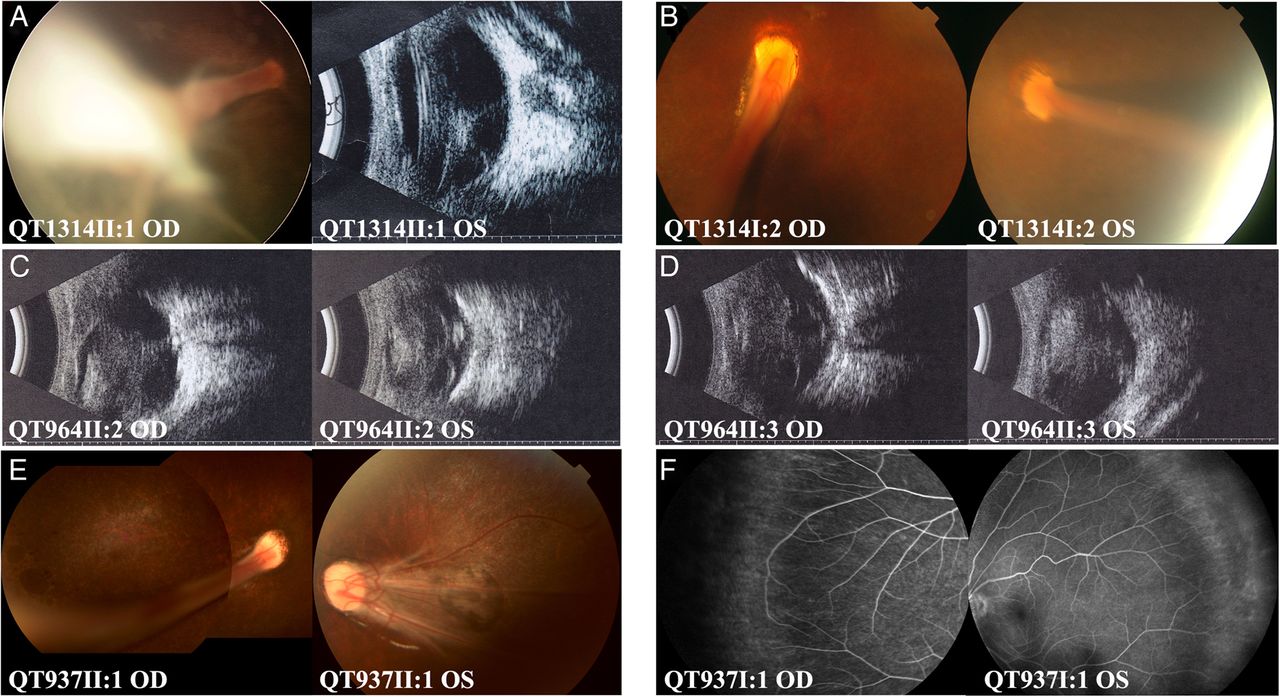
The seven patients from four families with KIF11 mutations showed typical signs of FEVR. Clinical data for patients with KIF11 mutations are summarised in table 3. All probands with KIF11 mutations had poor vision in early childhood. Visual acuity of individuals with mutations varied from normal, without any symptoms, to blind, with no response to light. Fundus changes varied significantly in different individuals, ranging from avascular zone of the peripheral retina to severe ocular changes, including temporal dragging of the optic disc, falciform retinal folds, retinal detachment and/or retrolenticular fibrotic masses in affected patients (figure 2). Microcephaly (3SD below the mean) was identified in four patients with KIF11 mutations, and lymphoedema in none. Mental retardation was not present in the two adult patients, QT1314I:2 and QT937I:1, but was present in QT761II:2 (identified by the parents when the patient was 5 years old) while the examination was not available for the remaining four patients.
Clinical information of the patients with KIF11 mutation

Fundus changes of patients with KIF11 mutations. Typical signs of familial exudative vitreoretinopathy (FEVR) included temporal dragging of the optic disc (A, B, E), falciform retinal fold (A, B, E), retinal detachment (C, D) and avascular zone in the peripheral retina (F). Retinal pigment changes (A, B, E) and chorioretinal atrophy (E) were present. The patient ID number is marked at the bottom left of each picture. OD and OS represent right and left eyes, respectively.
Mosaicism detected
In family QT964, the KIF11 c.2230C>T mutation was present in both siblings with FEVR, but not in the clinically asymptomatic parents with visual acuities of 20/20 and normal fundus examinations. Genotyping and haplotype analysis showed that the mutation KIF11 c.2230C>T might be inherited from the mother (figure 3A). Genetic testing of the mother showed a small variant peak for the mutant allele c.2230C>T (figure 3B), which might further indicate the existence of mosaicism with a low mutant allele frequency. After cloning sequencing, the mutation in 72 resultant clones, c.2230C>T was detected in about 7% (5/72) of the clones isolated from the mother's leucocyte DNA (figure 3C).
{kind=link}
{kind=link}
{kind=link}
Pedigree and haplotypes: sequence changes of family QT964. (DNA sample for II:1 was not available.) (A) Haplotypes of the KIF11 region of family QT964 showed the KIF11 c.2230C>T mutation and surrounding microsatellite markers. The risk haplotype is shown in black. (B) Sequence chromatography demonstrated a small variant peak which may refer to the KIF11 c.2230C>T mutation in individual QT964I:2. (C) Cloning sequencing demonstrated the KIF11 c.2230C>T mutation existing in the mother (QT964I:2). In the sequence chromatography, the variations are marked with arrows and named under the sequence.
In family QT761, cloning followed by sequencing of 32 clones was performed, but the c.2952_2955delGCAG mutant allele was not detected in any of the clones from both parents' leucocyte DNA, which suggested a de novo mutation in the proband, but not mosaicism in the parents.
Discussion
In this study, four novel KIF11 mutations were detected in four probands with FEVR, but not in the 780 inhouse controls. Segregation analysis in these four families indicated a dominant role of the mutations. These data provide evidence that the KIF11 mutations are the cause of FEVR in these patients.
Mutations in KIF11 have been identified in patients with MLCRD, which presented with four common features including microcephaly, mental retardation, lymphoedema and unclassified chorioretinopathy.12 A recent study identified mutations in KIF11 in a cohort of patients with FEVR.14 They found that KIF11 mutations were identified in four patients, all of whom showed typical signs of FEVR, two with microcephaly and none with mental retardation or lymphoedema. In our study, KIF11 mutations were identified in seven patients with FEVR. Systemic anomalies were also presented in seven patients, including four with microcephaly and one with mental retardation. The presence of patients without microcephaly, mental retardation or lymphoedema in our study was consistent with the findings of Jones et al12 and Robitaille et al.14 These patients were usually diagnosed as FEVR only, without MLCRD. However, it is important to evaluate the head circumference, lymphoedema and mental development in patients with FEVR for a comprehensive diagnosis. Thus, this study confirms and extends the findings of Jones et al and Robitaille et al that patients with KIF11 mutations can show a milder phenotype and can often be clinically labelled as FEVR.
Mutations in six genes have been reported to cause FEVR. In our previous studies, the mutation frequencies of the three known causative genes were 11.5% (6/52)16 in LRP5, 9.6% (5/52)16 in FZD4 and 5.7% (3/52)17 in TSPAN12, respectively. In the current study, KIF11 mutations were identified in 8.3% (4/48) of all the probands with FEVR and 12.9% (4/31) of probands without mutations in the four known FEVR genes. Therefore, mutations in KIF11 are one of the common causes of FEVR and KIF11 should be recruited for screening causative mutations in patients with FEVR. It is well known that the proteins encoded by the four known genes are important components of the Wnt/Norrin signalling pathway, which monitors retinal vascular development.23 Mutations in the four genes cause FEVR by affecting the Wnt/Norrin signalling pathway. KIF11 belongs to the kinesin family and encodes a spindle motor protein that contributes to the assembly of the bipolar spindle during mitosis.24 Whether KIF11 regulates the retinal vascular development through the Wnt/Norrin signalling pathway needs to be further studied. However, it suggests that more candidate genes responsible for FEVR might include other genes besides those in the Wnt/Norrin signalling pathway.
Mosaicism is a possible underlying mechanism to explain an affected child with phenotypically normal parents tested negative by routine mutational screening.25 The KIF11 mutation in the two siblings with FEVR was not identified in the phenotypically normal parents in family QT964 by Sanger sequencing. Since FEVR caused by KIF11 mutations is known to be inherited as an autosomal dominant trait, and the possibility of two de novo mutations happening independently in the same family was low, it was reasonable to suspect that mosaicism might be present in one of the parents in QT964. What was more; genotyping analysis and the small variant peak in the sequence chromatography might possibly indicate the existence of mosaicism in the mother, which was confirmed by clonal sequencing. To our knowledge, this is the first recognised somatic mosaicism of KIF11 mutations in patients with FEVR.
In summary, we have identified heterozygous KIF11 mutations as a common cause of autosomal-dominant forms of FEVR. There is a specific genotype-phenotype correlation for KIF11 in that more patients with KIF11 mutations showed microcephaly, lymphoedema or mental retardation when compared with the patients with other FEVR causative genes. Gene screening of patients with FEVR should include KIF11 in addition to the six known FEVR causative genes. In addition, somatic mosaicism for KIF11 mutation may exist in phenotypically normal parents. It should be taken into consideration in FEVR families with children who have clinically defined de novo mutations and with a family history counterintuitive for autosomal dominant inheritance. Accurate genetic counselling is necessary to avoid the recurrence of the same genomic disorder.
Acknowledgments
We thank the patients and the family members for their participation.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Data supplement 1 - Online supplement
Footnotes
Contributors QZ conceived and designed the study. XX, SL, XJ, XG and QZ contributed in collecting samples. HH, XX, SL, XJ and QZ participated in analysing and interpreting data. HH drafted the manuscript. QZ, XX, SL, XJ and XG revised the manuscript critically for important intellectual content. All authors approved the final version.
Funding The work was supported by grant U1201221 from the National Natural Science Foundation of China, S2013030012978 from Natural Science foundation of Guangdong, 2011A080300002 from Guangdong Department of Science & Technology Translational Medicine Centre, and by fundamental research funds of the State Key Laboratory of Ophthalmology (Dr. Zhang).
Competing interests None declared.
Patient consent Obtained.
Ethics approval The study was approved by the institutional review board of the Zhongshan Ophthalmic Centre.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- At a glance