Article Text

Abstract
BACKGROUND/AIMS Dendritic cells (DC) and macrophages are components of the immune cell populations in the uveal tract whose density, distribution, turnover, and function may play a role in the maintenance of immunological homeostasis in the eye. Little is known of these cells in the mouse eye despite this being the predominant experimental model in many studies of ocular immune responses and immunoinflammatory mediated eye diseases. The aim of the present study was to obtain further immunophenotypic data on resident tissue macrophages and DC populations in the mouse uveal tract.
METHODS Pieces of iris, ciliary body, and choroid dissected from perfusion fixed BALB/c mice were incubated whole in a variety of anti-macrophage and DC monoclonal antibodies (mAbs). Labelled cells were visualised using either single or double immunoperoxidase techniques.
RESULTS Quantitative analysis and double immunolabelling revealed that 80% of F4/80+ cells (a mAb that recognises both DC and macrophages) in the iris are macrophages (SER4+). The iris contained a network of Ia+ cells (412 (SD 130) cells/mm2) of which two thirds appear to be DC. A similar pattern was observed in the ciliary body and choroid. Only a few DC in the uveal tract were very weakly reactive for mAbs which recognise B7–1 (CD80), B7–2 (CD86), β2 integrin (mAb N418), and multivesicular bodies associated with antigen presentation (mAb M342).
CONCLUSIONS The present study reveals that the mouse uveal tract, like the rat, contains rich networks of DC and resident tissue macrophages. The networks of resident tissue macrophages in the mouse uveal tract closely resemble similar networks in non-ocular tissues. The phenotype of uveal tract DC suggests they are in the “immature” phase of their life cycle, similar to Langerhans cells of the skin, thus implying their role in situ within the eye is antigen capture and not antigen presentation.
- dendritic cell
- macrophage
- iris
- choroid
- immune privilege
- anterior chamber
- uveitis
Statistics from Altmetric.com
Understanding the mechanisms of immune surveillance which underlie immunological homeostasis in the anterior and posterior segment of the eye will inevitably lead to more rational approaches to therapy of inflammatory and immune mediated disorders such as anterior uveitis and endogenous posterior uveitis (see reviews by Forresteret al 1 2). The experimental phenomenon of anterior chamber associated immune deviation (ACAID) which is characterised by a selective inability to develop delayed hypersensitivity (DTH) responses following antigen administration into the anterior chamber (AC) has been postulated as a model for the study of the immune regulatory mechanisms in the anterior segment of the eye (see review by Streilein et al 3). Explanations of the mechanisms of this model propose that following antigen administration into the AC an antigen specific “signal” leaves the eye and traffics either to the spleen4 5 or the submandibular lymph node.6It is within either or both of these sites that distinctive populations of regulatory T and B cells are characteristically produced in ACAID (see review by Streilein et al 3). In mice, the species predominantly used in this model, it has been reported that the ACAID inducing “signal” leaving the eye is carried in the blood by “F4/80+monocytes/macrophages or APCs” derived from the iris/ciliary body.7-9 Whether these cells are of the dendritic cell (DC) and/or macrophage lineage is currently unknown as the monoclonal antibody F4/80 recognises both cell types.10 The ACAID inducing ability of these F4/80+ cells appears to be regulated by local “suppressor” factors within aqueous humour (AqH)11 such as TGFβ.12 Interestingly, the ability to transfer this form of tolerance does not appear to be eye specific as adherent peritoneal F4/80+ macrophages (not DC as these are non-adherent) can be substituted in ACAID experiments.13 Furthermore, these peritoneal macrophages are also influenced by TGFβ in a similar manner to ocular F4/80+ cells.14 This would naturally lead to the conclusion that the ACAID inducing signal leaving the eye was F4/80+ macrophages.
Immunohistochemical studies have revealed extensive networks of regularly arranged MHC class II+ DC and MHC class II- ED2+/ED1+ macrophages in the rat iris stroma, ciliary body, and choroid.15-19 To date only limited data have been presented on the distribution of DC and macrophages in the mouse iris.15 17 The first aim of the present study was to extend our preliminary observations17and more fully characterise potential “APC” populations (DC and/or macrophages) in the mouse iris using double immunohistochemistry. The second aim was to provide novel data on the distribution of DC and resident tissue macrophages in both the mouse choroid and ciliary body. The mouse offers some advantages over the rat in studies of this nature in that a wider range of monoclonal antibodies, for example, to accessory molecules and DC specific antigens, are available.20 21 It was hoped that use of these antibodies would give an indication of the functional status of uveal tract DC in the mouse.
Methods
ANIMALS
BALB/c mice (specific pathogen free, 8–12 weeks, n=60) obtained from the Animal Resources Centre (Murdoch, Western Australia) were used in the present study. Animals were housed in 12 hour day/night cycles. Throughout this study, all procedures conformed to the ARVO statement for the use of animals in ophthalmic and vision research.
METHODS
In most experiments the tissues were fixed and processed in a manner similar to that previously described for rat ocular tissues17 19; however, many other fixation protocols were investigated in an attempt to optimise antigen preservation (see below). Briefly, animals were anaesthetised (sodium pentobarbitone 100 mg/kg) before whole body perfusion via the left ventricle with cold heparinised phosphate buffered saline (PBS) followed by fresh cold 2% paraformaldehyde. Following removal of the lens, the iris, ciliary body, and choroid were dissected as whole mounts in the manner previously described. Pieces of iris/ciliary body and choroid were placed in PBS in 24 well plates where all subsequent immunohistochemical procedures were performed. Tissues were studied by single and double immunohistochemistry using a panel of primary monoclonal antibodies (mAbs) (Table 1) to investigate the phenotype of DC and resident tissue macrophages. Antibodies were purchased from either PharMingen (San Diego, CA, USA) and Serotec (Oxford, England) or kindly provided by Dr Ralph Steinman and Dr Patrick Holt. Iris/ciliary body complexes and choroids were incubated in warm 20 mM EDTA for 15 minutes and rinsed in a solution of PBS, 1% bovine serum albumin (BSA), and 0.2% Triton for 10 minutes before immunostaining. This step proved critical to successful immunostaining with some mAbs. A standard indirect immunoperoxidase procedure using primary mAb, biotinylated secondary antibody, and streptavidin-horseradish peroxidase (S-HRP) (Amersham Laboratories, Bucks) was used in this study. All procedures were performed at room temperature with incubation times of 30–45 minutes. The HRP was visualised using either 3.3 diaminobenzidine tetrahydrochloride (DAB; Sigma Chemical Co, St Louis, MO, USA) (1.2 mg/ml PBS and 0.5 μl/ml of hydrogen peroxide (30% v/v)) or 3-amino-9-ethyl carbazole (AEC) (0.2 mg/ml in acetate buffer pH 5; plus 0.35 μl/ml hydrogen peroxide (30% v/v)). Endogenous or pseudoperoxidase activity was not blocked as it was found to compromise antigenicity and tissue morphology. Cells displaying endogenous activity were easily distinguishable from immunostained cells. Stained whole mounts were mounted using aqueous mounting medium and coverslipped.
Monoclonal antibody (mAb) specificity
In an attempt to optimise staining with some mAbs (B7-1, B7-2, N418, M342) animals were fixed using a variety of protocols and fixatives, such as perfusion with either cold 100% ethanol, 2% paraformaldehyde, 4% paraformaldehyde, or PBS alone. The ocular tissues of some PBS perfused animals were post-fixed in ethanol, acetone, paraformaldehyde, or glutaraldehyde after application of primary and secondary antibodies. Optimal staining was obtained with anti-DC mAbs (M342 and N418) with very brief fixation with cold 2% paraformaldehyde. Staining with B7-2 was optimal when fresh PBS perfused tissue was postfixed with cold ethanol.
A double immunoperoxidase procedure26 which relies on one of the primary mAbs (F4/80) being biotinylated was used to investigate the extent of co-expression of leucocyte markers on macrophage and DC populations in the mouse uveal tract. Iris/ciliary body and choroid whole mounts were double stained with the following combinations: F4/80–M5/114; F4/80-SER4. Firstly, specimens were incubated with the non-biotinylated primary mAb, such as SER4, followed by directly conjugated secondary antibody (sheep anti-mouse-HRP, Amersham). The tissues were washed (PBS, ×3) and then incubated with the biotinylated primary mAb (F4/80-biotin) which was localised by streptavidin-alkaline phosphatase. The F4/80-alkaline phosphatase labelled cells were visualised with the substrate naphthol AS-MX (0.12 mg/ml) and fast blue BB (0.25 mg/ml) in TRIS buffer (pH 8.5). Levamisole (0.25 mg/ml) was added to the substrate to block endogenous alkaline phosphatase activity. Immunolabelled cells are visualised as a blue reaction product. Cells labelled with the other primary monoclonal (for example, SER4) and HRP conjugated secondary antibody were visualised using AEC (see above) which produces a red reaction product. Double staining or co-localisation of both phenotypic markers resulted in purple coloured cells or occasionally individual cells that contain discrete blue and red stained regions. All visualisation agents and chemicals were purchased from Sigma (Sigma Chemical Co, St Louis, MO, USA). Stained whole mounts were mounted in aqueous medium on glass slides and coverslipped.
CONTROLS
Positive controls for immunostaining procedures included frozen sections of lymphoid tissue (lymph node or spleen) or small whole mount preparations of non-fat bearing portions of mesentery from the small intestine (contains networks of resident tissue macrophages) and epidermal sheets from the pinna (contain Langerhans cells). Negative controls were also routinely performed. In these, all steps were performed with the exception of the primary antibody which was replaced with either PBS or an inappropriate mAb (isotype control). No staining was observed in negative controls.
Results
IRIS (FIG 1A-C, FIG 2A,B)
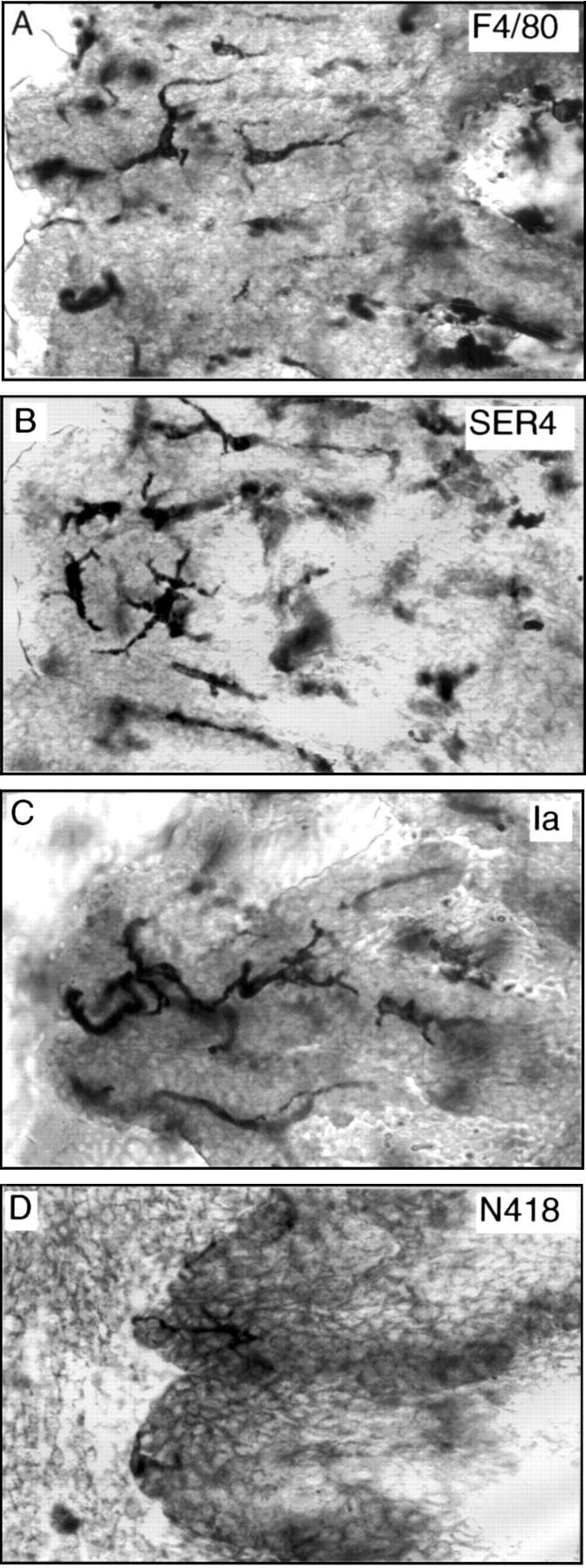
Single stained iris whole mounts revealed regular networks of dendriform and pleomorphic F4/80+ cells (637 (SD 58) cells/mm2, n=6) which extended from the iris base to the pupil margin (Fig 1A). Generally cells were regularly spaced relative to neighbouring cells and many displayed a perivascular arrangement. A minor subpopulation displayed a rounded morphology. Since F4/80 is a non-specific mAb which labels both macrophages and DC the staining pattern with other mAbs was investigated. A network of resident tissue macrophages of a similar density (669 (78) cells/mm2) to the F4/80+ cells was detected using a pan-macrophage specific antibody (SER4) (Fig 1B). Similar patterns were observed with other pan-macrophage mAbs (Mac1(CD11b) and MOMA2) (not illustrated). Double staining (Table 2) revealed that approximately 80% of all F4/80+ cells were SER4+ and therefore likely represent macrophages. In addition 98% of all SER4+ macrophages were F4/80+. Single staining revealed a network of MHC class II (Ia)+ DC (412 (130) cells/mm2) (Fig 1C). Double staining revealed that around 33% of F4/80+ cells co-expressed Ia (Table 2) and therefore could represent DC or Ia+ macrophages. It should be noted that staining with anti-Ia mAb was slightly more variable between groups of animals than other mAbs. For example, the mean density of Ia+ cells in the animals used in the double immunostaining experiments was 268 (80) cells/mm2 (Table 2), approximately 30% less than in single immunostaining experiments. However, it should be noted that the total number of cells in double immunohistochemical techniques is generally slightly lower than single immunohistochemical techniques.

Low power and medium power views (insets) of mouse iris whole mounts stained with F4/80 (A), SER4 (B), and Ia (MHC class II) (C). In low power micrographs the pupil margin is on the left (arrow in A) and the ciliary body is on the right. Magnifications: A, B, C, ×80; inserts, ×180.
Double immunohistochemical staining results (n=6)
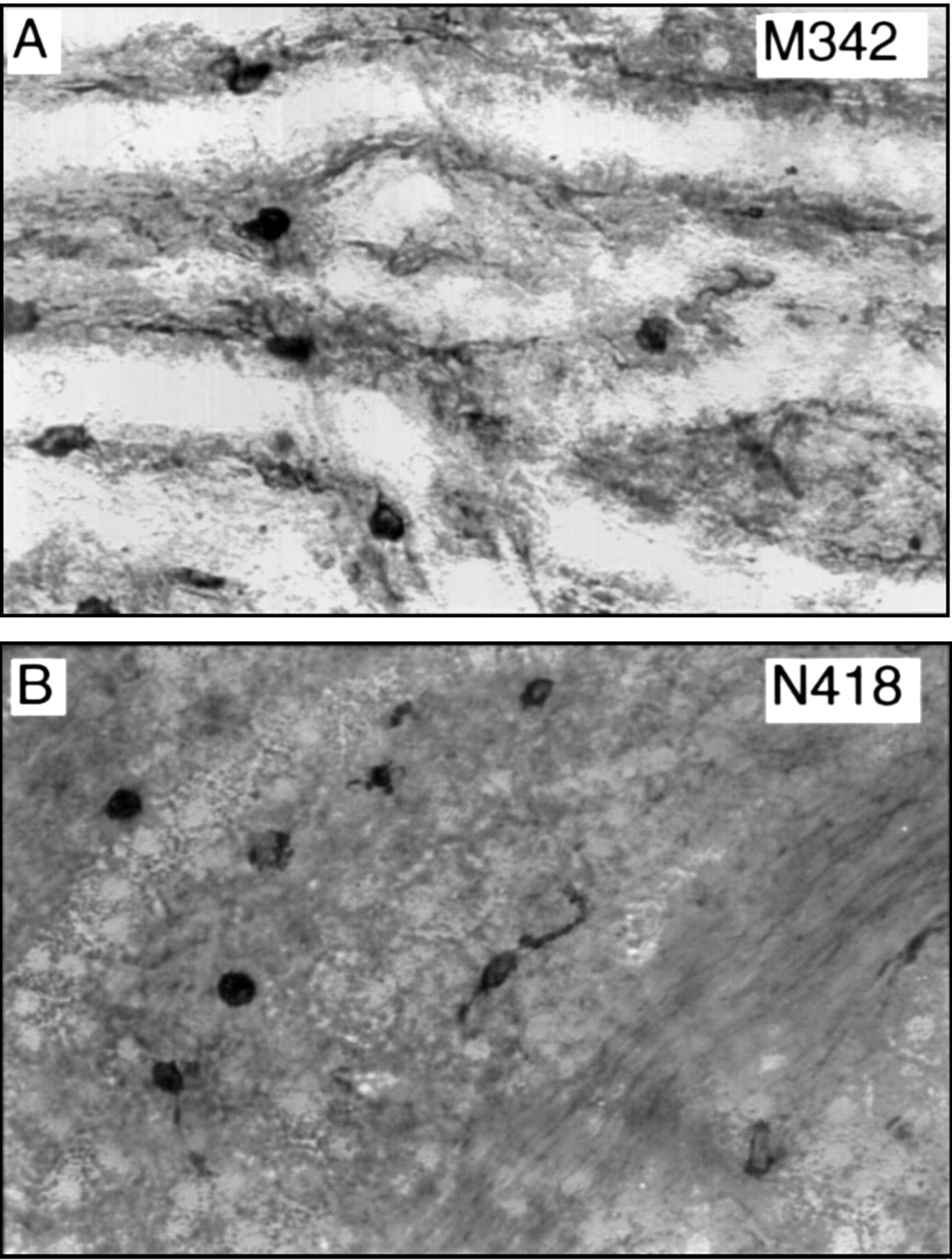
No staining with B7-1 (CD80) and B7-2 (CD86), accessory or costimulatory molecules important in antigen presentation, was noted in paraformaldehyde fixed tissue. Optimal staining results were obtained by incubating the primary mAb with PBS perfused (unfixed) tissue followed by fixation with cold ethanol. In these preparations positive control tissue stained satisfactorily but only very weakly positive cells which lacked distinct cell processes were identified in the iris (Fig 2A). In order to illustrate these cells the contrast in Figure 2A has been markedly enhanced and somewhat overrepresents the degree of staining with B7-1. Similarly, application of DC specific hamster anti-mouse mAbs (M342, N418) (see Table 1 for specificities) revealed only very weakly immunopositive cells in the iris (Fig 2B) in tissue briefly fixed in 2% paraformaldehyde and incubated in EDTA and Tween (see Methods). Iris DC were negative for NLDC-145, a mAb which recognises mannose receptors on lymphoid DC (Table 1), despite numerous fixation protocols. Positive control tissues revealed immunoreactive cells in accordance with previous descriptions.20 21

Medium power light micrographs of mouse iris whole mounts stained with B7-1 (A) and N418 (B) mAbs against DC specific antigens. Note the contrast has been enhanced to demonstrate the weakly stained cells. Magnifications: A, B, ×180.
CILIARY BODY (FIG 3A–D)
Networks of dendriform and pleomorphic F4/80+, SER4+ macrophages were identified in the stroma of the mouse ciliary body and close to the ciliary epithelial layers (Fig3A,B). Care has to be taken when examining the ciliary body as cells in the iris base may overlie the ciliary processes (Fig 1C) and can lead to false impressions. To avoid this irides were frequently removed from the ciliary body before mounting and care was taken to firstly focus on and identify the ciliary epithelial cells and stroma (Fig 3A-C). Ia+ DC and a few weakly stained N418+ DC were identifiable in the ciliary body in the plane of the ciliary epithelium. The ciliary body Ia+ DC were more dendriform than iris DC, a feature also observed in rats.17

High power views of mouse ciliary processes stained with a variety of mAbs. Note the Ia+ and N418+Dendritic cells are in the same focal plane as the ciliary epithelium. Magnifications: A–D, ×290.
CHOROID (FIG 4A–C; FIG 5A,B)
Regular arrays of F4/80+ and SER4+macrophages were identified within the mouse choroid. The majority of these cells displayed a perivascular arrangement and were pleomorphic, dendriform, or fusiform in shape (Fig 4A,B). An extensive network of Ia+ DC was identified in the choroid (Fig 4C). These cells were more dendriform in shape than the resident tissue macrophages and also displayed a less perivascular distribution. The pattern of double staining in the choroid was identical to the iris. Quantitative analysis of cell density or double staining was not performed on choroidal tissue. Staining with the anti-DC mAbs M342 and N418 in addition to the B7-1 and B7-2 mAbs revealed only scattered weakly stained cells which were generally rounded in shape and only rarely dendriform (Fig 5A,B). Staining with the mAbs M342 and N418 appeared marginally stronger than in the choroid than in the iris. As observed in the iris no NLDC-145 reactive cells were identified in the choroid.

Low and medium (insets) power views of choroidal whole mounts stained with the mAbs F4/80 (A), SER4 (B), and Ia (C). Magnifications: A, B, C, ×80; insets, ×180.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Weakly positive cells in the mouse choroid stained with mAbs M342 and N418. Note most cells have a rounded morphology. Magnifications ×290.
Discussion
The present study is the first detailed immunohistochemical analysis of resident tissue macrophages and DC in the mouse uveal tract. The data presented extend our preliminary study which briefly documented the distribution of DC and macrophages in the mouse and human iris and ciliary body in the context of a more extensive study of these cells in the rat eye.17 The present investigation reveals for the first time that the vast majority of F4/80+cells in the mouse anterior uveal tract are SER4+ resident tissue macrophages and are of a similar density (∼600–700 cells/mm2) to ED2+ macrophages in the rat eye.17 It also shows that there is a similar density of highly dendriform MHC class II+ cells in the mouse iris, ciliary body, and choroid to that described in rat eye (∼400–500 cells/mm2)17 19 which on the basis of double immunolabelling are most likely DC. Further mAb combinations not performed in the present study—for example, M5/114-SER4, could help clarify the exact number of DC. The present data contrast with previous studies by Williamson and co-workers7 in which they found considerably fewer Ia+ cells than F4/80+macrophages and observed only rare Ia+ cells with a dendriform morphology. They therefore concluded that the majority of “bone marrow derived” cells in the mouse iris/ciliary body were “F4/80+ macrophages/monocytes” and “not of the DC-Langerhans cell lineage” (original authors’ italics). It is likely that the difference between the present study and that of Williamson et al 7 is due to their use of acetone fixation compared with the paraformaldehyde based fixative used in both the current investigation and that by Knisely et al.15 The latter group also described many highly dendriform Ia+ cells in the mouse iris. Unfortunately, exact comparisons of the cell densities between the present and previous studies is not possible as Knisely et al 15 did not perform quantitative analysis and Williamson et al 7 reported densities as cells per microscopy field and not as cells per mm2.
The importance of distinguishing DC from macrophages in studies such as the current one or in investigations of experimental ocular immune responses can be readily appreciated when the functions of these two distinct lineages of immune cells are compared. Members of the DC family play a pivotal role in the initiation of antigen specific adaptive immune responses.27 28 Tissue resident DC are characterised by their dendritic morphology, constitutive MHC class II expression, lack of expression of pan-macrophage markers, weak phagocytic activity, and contiguous network arrangement. While resident in peripheral tissue sites DC are considered to be in the “immature” phase of their life cycle and specialised for antigen acquisition and processing and possess only a weak immunostimulatory capacity.29 After a defined period of residence or following cytokine mediated signals DC migrate via lymphatics or blood vessels (as “veiled” cells) to the T cell dependent areas of “local” lymphoid tissues.30 Here they mature, under the influence of cytokines, particularly GM-CSF,31 into functionally potent antigen presenting cells (APC) and present antigens/peptides within the groove of the cell surface MHC class II molecule to resting T cells.32 Expression of secondary costimulatory molecules, such as B7-1 and B7-2 (CD80 and CD86) both ligands for CD28/CTLA-4, and adhesion molecules are crucial for effective MHC class II/T cell receptor interactions and increase T cell activation24 by enhancing IL-2 mRNA levels. The specialisation of DC for antigen trapping, processing, and presentation, along with their migratory properties and life cycle has led to them being regarded as “sentinels” of the immune system.27 In addition, they are many times more potent at initiating and perpetuating secondary immune responses than other APC such as B cells and macrophages. The absence of DC from the CNS neural parenchyma33 and their poor APC activity in situ in some peripheral sites, such as the anterior chamber,34 could be interpreted as a means of minimising local potentially damaging DTH responses within such delicate and functionally important regions.
The weak immunoreactivity of DC in the mouse iris, ciliary body, and choroid with antibodies to β2 intigrin (CD11c) (mAb N418) and intracellular multivesicular bodies involved in antigen presentation (mAb M342) supports the author’s proposal that ocular DC, in common with other peripherally located non-lymphoid DC, are at the “immature” or antigen capturing/trapping phase of their life cycle. These two anti-DC mAbs react more strongly with subpopulations of mouse lymphoid DC such as interdigitating cells which are in the “mature” phase of their life cycle, when antigen presentation is their primary role. The weak or poor B7-1 and B7-2 reactivity of mouse uveal tract DC would also support this conclusion. Taken together the evidence suggests that mouse ocular DC may be poor APCs in situ within the eye and require maturational signals during their migration. While no functional analysis of purified mouse ocular DC is available the hypothesis is supported by functional studies in the rat34which have shown that purified iris DC were poor APCs in the fresh state but following incubation with GM-CSF they became potent allostimulatory cells to naive T cells.
The heterogeneous nature, function, morphology, and immunophenotype of the monocyte/macrophage series in different tissue microenvironments is well established35 36 and includes Kupffer cells, mesangial cells, alveolar macrophages, microglia (brain parenchyma and retina), peritoneal macrophages, and other resident tissue macrophage populations widely distributed throughout the body. Macrophages are professional phagocytes and play a pivotal role as effector cells in cell mediated immunity and inflammation and in other processes including immune regulation, tissue reorganisation, and angiogenesis.35 Activated macrophages may express low/moderate amounts of MHC class II molecule and thus play a role in antigen presentation in secondary immune responses.35Newly recruited macrophages (responsive macrophages) at sites of inflammation undergo a complex process of activation driven by molecular inductive signals—for example, IFNγ. Fully activated macrophages are capable of effecting a range of destructive powers towards foreign organisms or tumour cells. Depletion of bone marrow derived macrophages using liposomes containing dichloromethylene diphosphonate before the effector phase of experimental autoimmune encephalomyelitis37 and experimental autoimmune uveoretinitis (EAU)38 (models of multiple sclerosis and posterior uveitis, respectively) show that responsive macrophages have an important role in the pathogenesis of these diseases although depletion was unable to completely abrogate the inflammatory response. Resident tissue macrophages are poorly responsive to activation signals.36 Recent data using the EAU model support this proposal19 38 and showed the network of ED2+ resident tissue macrophages was largely unaltered in the early and late phases of the disease. The precise role of such extensive networks of resident tissue macrophages in normal tissues is a less clearly understood aspect of macrophage biology.
As stated in the introduction it has been reported that the antigen specific ACAID inducing signal leaving the eye is carried by “F4/80+ macrophages”. It is now clear from the present study and the brief comparison of DC and macrophage function outlined above that the prime candidate for the role of an antigen carrying cell capable of trafficking from the eye to lymphoid tissues is the DC population in the uveal tract. This is difficult to reconcile with the experimental data which report that the ACAID inducing signal can be mimicked by adherent peritoneal macrophages.17 Clearly it would be valuable if the exact immunophenotype and APC capacity of cells used in models of ACAID were clarified in future experiments. Furthermore, experiments are required to determine whether the networks of ocular DC can trap antigens placed in the AC. Such experiments are currently being performed in the author’s laboratory.
The present study reports novel data on the presence of DC and resident tissue macrophage networks in the mouse choroid. The densities and distribution are essentially similar to those described in the rat18 19 although quantitative analysis has not been performed in the present study. The observation that choroidal DC were more immunoreactive with the mAbs N418 and M342 than iris DC is of some interest. In a previous study in the rat the expression of ED3 (a mAb recognising an antigen normally expressed by lymphoid macrophages) on resident tissue macrophages in the choroid was found to be greater than iris macrophages.19 This suggests the extravascular compartment of the choroid and ciliary body with their fenestrated capillary beds represent a different milieu for immune cells when compared with the iris. This probably reflects the position of these cells relative to the blood-ocular barrier—namely, choroidal cells lie outside the barrier whereas the iris cells are within the barrier (see reviews by McMenamin39 40). It may also be significant that immune cells in the iris are bathed in aqueous humour which has been described as having immunosuppressive properties.11 12 Thus it could be postulated that antigen presentation by DC to circulating autoreactive T cells is more likely to occur locally in the choroid and ciliary body than in the iris. This may have important bearings on the pathogenesis of anterior and posterior uveitis.
Acknowledgments
The author wishes to acknowledge support for this research by the Australian NH&MRC and would also like to thank Mr Colin Murphy, Photographic Unit, University of Western Australia, for assistance with production of the photographic plates.