Article Text

Abstract
Background Deregulated expression of vascular endothelial growth factor (VEGF), basic fibroblast growth factor (bFGF) or insulin-like growth factor-1 (IGF-1) is associated with the pathogenesis of diabetic retinopathy. The VEGF165-induced increase in permeability of retinal endothelial cells (REC), probably resulting in diabetic macular oedema (DME), could be completely restored by the VEGF-binding Fab fragment ranibizumab in vitro. We investigated whether bFGF and IGF-1 as single factors or in combination with VEGF165 influence permeability and tight junctions in immortalised bovine REC (iBREC) and if these effects could be restored by inhibition of VEGF.
Methods As a measure of changes in cellular permeability, transendothelial electrical resistance (TER) was monitored during long-term treatment of iBREC with growth factors in the absence or presence of ranibizumab or KRN951 (an inhibitor of VEGF receptors). Expression of claudin-1, as an indicator of functional tight junctions, was assessed by western blot analysis.
Results Whereas VEGF165 decreased TER and expression of claudin-1 in a concentration-dependent manner, long-term treatment of iBREC with 10–100 ng/ml bFGF or/and IGF-1 did not. Changes in claudin-1 expression or TER, induced by 25 ng/ml VEGF165, were slightly enhanced by bFGF and/or IGF-1 and were accompanied by a slightly increased secretion of VEGF. Complete reversion of these effects was achieved by prolonged treatment with ranibizumab and partly by exposure to KRN951.
Conclusion Our findings indicate that VEGF165, but not IGF-1 or bFGF, is mainly responsible for changes in cellular permeability observed in REC. This supports VEGF targeting as a therapeutic concept for DME.
- Retinal endothelial cells
- VEGF
- bFGF
- IGF-1
- VEGF inhibitor
- ranibizumab
- diabetic macular oedema
- retina
- macula
- biochemistry
- experimental and laboratory
Statistics from Altmetric.com
- Retinal endothelial cells
- VEGF
- bFGF
- IGF-1
- VEGF inhibitor
- ranibizumab
- diabetic macular oedema
- retina
- macula
- biochemistry
- experimental and laboratory
Introduction
It is a now generally accepted concept that pathogenesis of diabetic retinopathy is driven by deregulated expression of angiogenic growth factors such as vascular endothelial growth factor (VEGF), insulin-like growth factor (IGF-1) and basic fibroblast growth factor (bFGF). The levels of these proteins were found substantially elevated in the vitreous fluid and in the retinal vasculature of patients with diabetic retinopathy.1–5 VEGF and bFGF are involved in embryonic development of the vasculature and also play a major role in tumour neo-angiogenesis.6 7 In vitro studies confirmed an involvement of bFGF, IGF-1 and VEGF in stimulation of proliferation and migration of retinal endothelial cells (RECs), probably leading to neovascularisation seen in proliferative diabetic retinopathy.8–10 Induction of VEGF and VEGF receptor 2 (KDR) by bFGF or IGF-1 in REC and other cell types has been suggested to contribute to pathogenic mechanisms.11 12 An increased concentration of VEGF directly results in elevated permeability of REC and, as a consequence, in diabetic macular oedema (DME) that may appear at any stage of diabetic retinopathy.13 14 Elevated permeability was associated with altered cellular localisation, expression and/or modification of tight junction (TJ) proteins, and the presence of plasma membrane-localised claudin-1 strongly correlates with barrier function in REC in vivo and in vitro.12 15–17 Whereas overexpression of IGF-1 in the retina of transgenic mice results in the breakdown of the blood–retina barrier associated with higher expression of VEGF, the role of bFGF in this process is still unclear.12 Based on these observations, we studied whether long-term treatment of immortalised endothelial cells of the bovine retina (iBREC) with bFGF and IGF-1 applied as single agents or in combination with VEGF165 changed cellular permeability and/or claudin-1-expression as an indicator of barrier function in iBREC. In addition, we investigated if inhibition of VEGF by the VEGF-binding Fab fragment ranibizumab was sufficient to restore these effects.
Materials and methods
Reagents and antibodies
Recombinant human growth factors bFGF, IGF-1 (both E. coli-expressed) and rhVEGF165 (SF21-expressed) were from R&D Systems (Wiesbaden, Germany). The modified Fab fragment ranibizumab of a humanised VEGF-binding antibody was a gift from Novartis Pharma GmbH (Nuremberg, Germany).18 KRN951 (N-(2-chloro-4-((6,7-dimethoxy-4-quinolyl)oxy)phenyl)-N'-(5-methyl-3-isoxazolyl)urea), a tyrosine kinase inhibitor specific for VEGF receptors, was obtained from Calbiochem (Merck, Darmstadt, Germany).19 The Complete EDTA-free protease inhibitor cocktail was purchased from Roche Applied Science (Mannheim, Germany). Polyoxyethylene (10) oleyl ether (Brij 97), sodium deoxycholate and the phosphatase inhibitor cocktail 2 were from Sigma-Aldrich (Deisenhofen, Germany). Rabbit polyclonal antibodies binding to human claudin-1 (JAY.8), claudin-5 (Z43.JK) and zona occludens 1 (ZO-1) and AlexaFluor 594 conjugated detection antibodies (F(ab')2) were from Invitrogen (Karlsruhe, Germany); rabbit polyclonal antiserum binding to human KDR, VEGF receptor 1 (Flt-1), cadherin 5 (VE-cadherin) and neuropilin were from Acris (Herford, Germany).
Cell cultivation
Telomerase-immortalised microvascular endothelial cells from bovine retina (iBREC) were cultivated on fibronectin-coated surfaces in cell culture flasks (BD Biosciences, Heidelberg, Germany) in complete microvascular endothelial growth medium (ECGM; Promocell, Heidelberg, Germany) supplemented with co-delivered premixed additives resulting in final concentrations of 0.4% ECGS/H (endothelial cells growth supplement/H), 5% fetal calf serum, 10 ng/ml epidermal growth factor and 103 nM hydrocortisone.10 Cells used in the experiments were at passages 20 to 35 counting from the stage of primary culture, for which stable expression of investigated proteins had been confirmed.
Treatment of iBREC with growth factors
Prior to the experiment with confluent iBREC, ECGM was replaced by serum-reduced medium supplemented with 0.4% ECGS/H, 0.25% fetal calf serum and 103 nM hydrocortisone for 24 h. Cells were incubated with 10–100 ng/ml VEGF165, bFGF or IGF-1, or with combinations of these factors, for up to 2 days before whole cell extracts were prepared. To study the effect of VEGF inhibitors, 100 μg/ml ranibizumab or 1 to 100 nM KRN951 were added for 1 to 3 days to the cells that had been treated with growth factors.
Western blot analyses
To prepare extracts, 3×106 cells were detached by scraping, collected by centrifugation and resuspended in 100 μl cold lysis buffer (40 mM TrisCl, 150 mM NaCl, 0.5% sodium deoxycholate, 1% Brij 97, pH 7.4, supplemented with EDTA-free protease inhibitor and phosphatase inhibitor cocktails). After incubation on ice for 1 h with gentle agitation, components that were still insoluble were removed by centrifugation (10 000×g, 4°C) for 45 min. Proteins (10 μg per lane or 20 μl of cell culture supernatant fraction) were separated by electrophoresis in a 4–20% SDS–polyacrylamide gradient gel (BioRad, Munich, Germany) under reducing conditions and transferred by electroblotting in transfer buffer (192 mM glycine, 25 mM Tris, pH 8.4) to a polyvinylidene difluoride membrane (BioRad).10 16 Antigen-bound primary antibodies were detected using appropriate IgG horseradish peroxidase conjugates (1:7500 in Mg2+-/Ca2+-free PBS, 0.1% Tween-20; BioRad) and the ECL-chemiluminscence kit (GE Healthcare, Munich, Germany) according to the supplier's protocol. To verify loading of equal amounts of protein, expression of actin was assessed in parallel using mouse monoclonal antibody AC-40 (Abcam, Cambridge, UK).
VEGF potentially secreted into the culture medium of iBREC treated with bFGF or IGF-1 was analysed after 20-fold concentration of the medium by ultrafiltration with Microcon centrifugal devices (cut-off size 10 kDa; Millipore, Schwalbach, Germany) according to the manufacture's instructions. Culture medium of iBREC treated with VEGF165 only or with combinations of factors was not concentrated prior to western blot analyses. To detect all VEGF isoforms, a polyclonal goat anti-canine VEGF antibody (IgG fraction, 0.2 μg/ml; R&D Systems) was used.
All western blot analyses were repeated at least twice.
Transendothelial electrical resistance of cell layers
To assess paracellular permeability of iBREC, transendothelial electrical resistance (TER) measurements were performed as described previously: polyethylenterephthalate membrane inserts pre-coated with fibronectin (0.3 cm2, pore size 0.4 μm; BioCoat, BD Biosciences) were placed in a 24-well plate and incubated with ECGM for 1 h at room temperature. On each of these, 5×103 cells were cultivated in fresh medium for 3 days until a confluent layer was formed. ECGM was replaced by serum-reduced medium 24 h before growth factors were then added as described earlier. TER measurements were made with a Millicell ERS resistance meter (Millipore, Schwalbach, Germany) just before and after addition of growth factors at given time points.16 To assess the effect of VEGF inhibitors, the medium was exchanged by medium containing growth factors with or without 100 μg/ml ranibizumab or 10 nM KRN951 48 h after addition of growth factors. Resistances (deltaTER) of iBREC layers under different conditions were calculated as means of at least four replicates from which the mean resistance of control inserts (without cells, four replicates) was subtracted. To compare independent experiments, normalised deltaTER values were calculated in relation to the deltaTER measured in low serum medium just before addition of effectors.
Statistical analyses
All experiments were repeated at least three times and in each experiment, data were generated from multiple replicates. The Mann–Whitney U test was used to analyse experimental data and resulting p values below 0.05 were considered indicative of significant differences.
Results
Effect of VEGF165 on iBREC barrier function
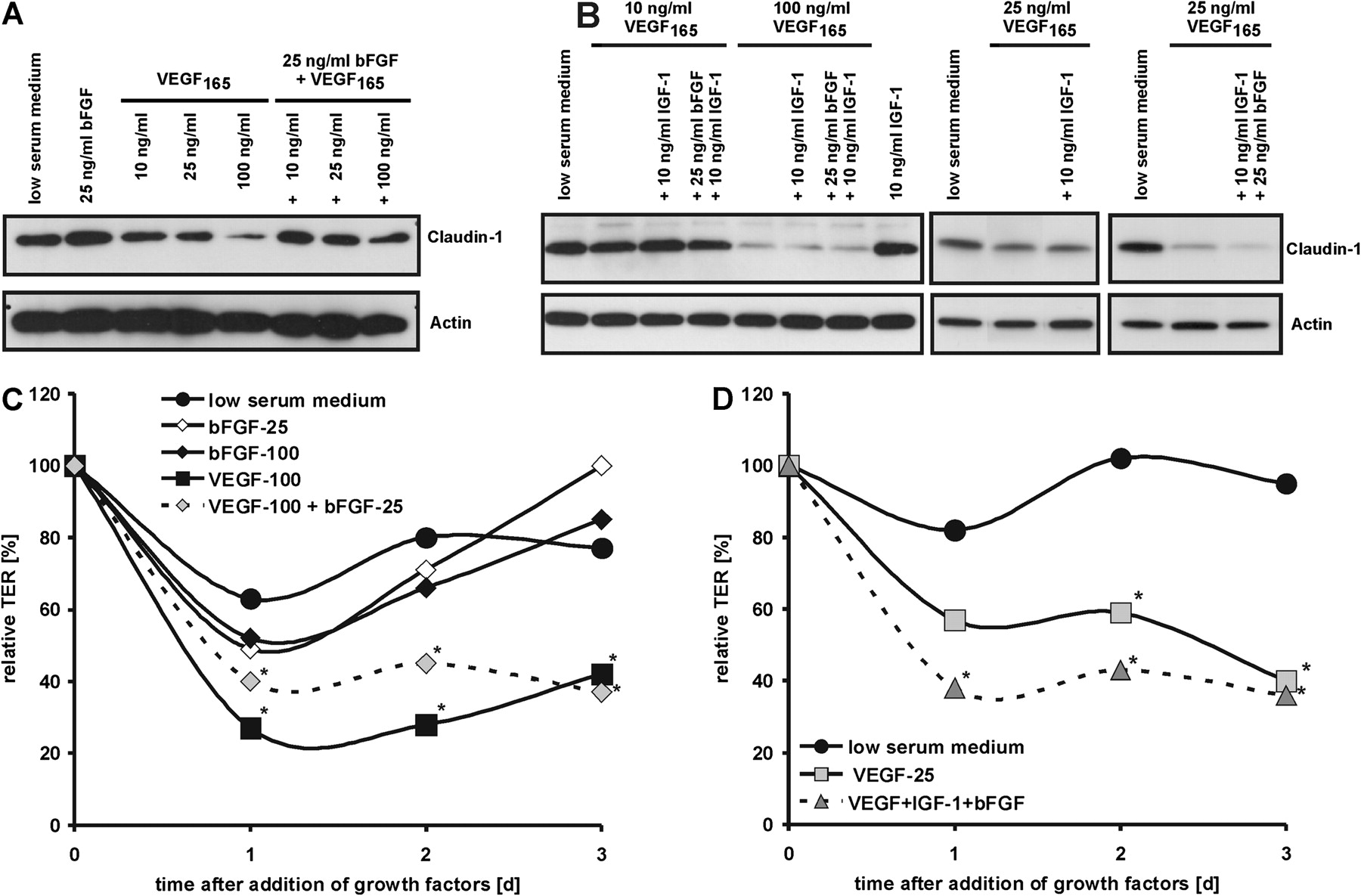
In all experiments iBREC formed a confluent monolayer 3 days after seeding. Culture medium always contained 103 nM hydrocortisone to ensure high expression of TJ proteins in these cells.15 20 Under these conditions, iBREC also formed a tight barrier reflected by a stable TER of the monolayer of about 30 Ω×cm2, varying in the range from 15 to 45 Ω×cm2 in accordance with values reported for monolayers of primary BREC.21 For further analyses, TER values were normalised. To determine the range of concentrations at which VEGF165 increases permeability of iBREC, TER of confluent cells incubated with 10 to 100 ng/ml VEGF165 was measured over a period of 3 days (figure 1A). In parallel, presence of TJ proteins in whole cell extracts was monitored by western blotting (figure 1B,C). A VEGF165 concentration of 50 ng/ml was sufficient to reduce TER significantly within 1 day, which correlated with reduced or absent expression of claudin-1. Whereas a low concentration of 10 ng/ml VEGF165 did not alter TER and measured amounts of claudin-1, 25 ng/ml VEGF165 resulted in a significant decrease of TER during prolonged treatment for 2 days. Several other proteins involved in barrier regulation were not affected after long-term treatment of iBREC with 100 ng/ml VEGF165 (figure 1C).

Concentration-dependent decrease in TER and claudin-1 induced in iBREC by VEGF165. Confluent iBREC were treated with 10 to 100 ng/ml VEGF165. (A) TER was measured at indicated time points. (B, C) Expression of claudin-1 and other proteins involved in barrier regulation was analysed by western blot after incubation for 2 days. The TER values shown were normalised in relation to deltaTER in low serum medium at t=0 days, which was set 100%. For details see text. Whereas 10 ng/ml VEGF165 did not change TER or claudin-1 expression, higher concentrations resulted in a decrease of TER correlating with loss of claudin-1. Other proteins appeared unchanged after long-term treatment of iBREC with 100 ng/ml VEGF165. *p<0.02 compared with low serum medium.
Effect of IGF-1 and bFGF as single factors or in combination with VEGF165 on iBREC barrier function
TER and claudin-1, as indicators of a functional barrier, were measured in iBREC treated with the growth factors bFGF or IGF-1 for at least 2 days (figure 2). Although 25 ng/ml bFGF increased claudin-1 expression slightly after treatment for 2 days, this was not sufficient to prevent loss of claudin-1 induced by VEGF165 (figure 2A). Accordingly, even long-term treatment of iBREC with 25 ng/ml bFGF did not result in significant changes in TER compared with untreated cells, and the VEGF165-induced TER decrease was also not affected by bFGF (figure 2C). Like bFGF, IGF-1 (10–100 ng/ml) did not alter claudin-1 expression or TER in iBREC, or VEGF165-induced changes in these properties (figure 2B).

Effects of bFGF and IGF-1 on VEGF165-induced decrease of claudin-1. (A, B) Confluent iBREC were treated with VEGF165, bFGF and/or IGF-1 at various concentrations. Expression of claudin-1 was analysed by western blot after incubation for 2 days. (C, D) TER was measured at indicated time points after addition of 25 ng/ml VEGF165 (VEGF-25) or 100 ng/ml VEGF165 (VEGF-100) with or without 25 ng/ml bFGF or 10 ng/ml IGF-1, or after addition of 25 ng/ml or 100 ng/ml bFGF (bFGF-25 and bFGF-100, respectively). The TER values shown were normalised in relation to TER in low serum medium at t=0 days, which was set 100%. For details see text. Only at 25 ng/ml VEGF165, IGF-1 and bFGF slightly increased the effect of VEGF165 on claudin-1 expression. *p<0.02 compared with low serum medium.
In iBREC treated only with bFGF or IGF-1, most of the claudin-1 protein is found at the plasma membrane with only a small portion showing intracellular localisation, whereas it delocalises from the plasma membrane in the presence of VEGF165. This was also observed when either bFGF or IGF-1, or both, had been added (results not shown).
In accordance with results obtained with single factors, 25 ng/ml bFGF together with 10 ng/ml IGF-1 did not change expression of claudin-1. Modulation of the action of VEGF at low or high concentrations by a combination of both growth factors was also not seen. However, treatment of iBREC with both factors and 25 ng/ml VEGF165 for 2 days resulted in an even stronger decrease of claudin-1 than observed with VEGF165 alone (figure 2B), whereas expression of other proteins involved in barrier regulation did not change markedly under these conditions (figure 3). TER of iBREC treated with 25 ng/ml VEGF165/25 ng/ml bFGF/10 ng/ml IGF-1 was significantly reduced after treatment for 1 d whereas TER of iBREC only exposed to 25 ng/ml VEGF165 was not (figure 2D). These findings suggested that combined action of IGF-1 and bFGF intensified the effects of VEGF165 applied at a moderate concentration resembling the physiological conditions in the vitreous fluid.

Effect of long-term treatment with VEGF165/bFGF/IGF-1 on expression of barrier-associated proteins in iBREC. Confluent iBREC were treated with 25 ng/ml VEGF165 in combination with 25 ng/ml bFGF and 10 ng/ml IGF-1 for 2 days before ranibizumab or KRN951 were added, and protein expression was analysed by western blot 24 h later. Only claudin-1 expression was altered under these conditions.
Effect of ranibizumab on decrease of TER or claudin-1 expression induced by VEGF165 plus IGF-1 or bFGF
Because IGF-1 and bFGF did not change barrier properties of iBREC, we assumed that increase in permeability and loss of claudin-1 induced by treatment with these factors and VEGF165 would be completely restored by inhibition of VEGF using the VEGF-binding Fab fragment ranibizumab. Addition of 100 μg/ml ranibizumab completely prevented or reverted VEGF-induced loss of claudin-1 or reduction of TER within 24 h (figure 4A).15 16 When 100 μg/ml ranibizumab was added to iBREC treated for 2 days with combinations of VEGF165 and bFGF or IGF-1, the TER increased significantly within 24 h and reached normal values after 4 days (figure 4B). Ranibizumab also reinstated lost claudin-1 within 24 h (figures 3 and 4A). To investigate effects of interfering with VEGF-mediated signalling, KRN951, a specific and potent (IC50∼0.2 nM) inhibitor of VEGF-induced phosphorylation of KDR and Flt-1 was added. Loss of claudin-1 in iBREC treated with 75 ng/ml VEGF165 could be completely prevented with less than 1 nM KRN951 (figure 4C), indicating that the inhibitor efficiently targets the VEGF receptors in these cells. However, addition of 10 nM KRN951 to iBREC treated with 25 ng/ml VEGF165/10 ng/ml IGF-1/25 ng/ml bFGF for 2 days only partly restored lost claudin-1 after 24 h (figures 3 and 4A). TER values did not significantly increase under these conditions. The effects of 1 nM or 100 nM KRN951 were similar (data not shown).

TER and claudin-1 decreased by growth factors are restored by inhibition of VEGF. (A) Confluent iBREC were treated with VEGF165 alone or in combination with bFGF or bFGF and IGF-1 for 2 days before ranibizumab or KRN951 were added. Expression of claudin-1 was analysed by western blot 24 h later. Inhibition of VEGF restored lost claudin-1. (B) TER was measured at indicated time points after adding 100 ng/ml VEGF165 and 25 ng/ml bFGF or 25 ng/ml VEGF165 with 25 ng/ml bFGF and 10 ng/ml IGF-1 to confluent iBREC for 2 days. Ranibizumab (100 μg/ml) was then added as indicated. Complete reversion of TER decrease was achieved after treatment for 4 days. (C) Confluent iBREC were treated with 0.5 to 50 nM KRN951 for 1 h before VEGF165 was added, and expression of claudin-1 was analysed by western blot 24 h later. Addition of less than 1 nM KRN951 prevented loss of claudin-1. *p<0.02 compared with low serum medium; #p<0.03 compared with low serum medium and for growth factors versus growth factors+ranibizumab.
Secretion of VEGF by iBREC treated with growth factors
Our findings suggested that IGF-1 and bFGF strengthen or help to maintain the actions of VEGF165 that lead to altered permeability or expression of claudin-1. Because this effect was completely restored by inhibition of VEGF only, we determined by western blot analyses the amounts of VEGF present in iBREC treated for 3 days with either IGF-1 or bFGF, or both factors, in the absence or presence of exogenous VEGF165 (figure 5). Without addition of VEGF165, endogenous VEGF in whole cell extracts or in concentrated culture medium was below the detection limit under all conditions tested. In culture medium of iBREC treated with VEGF165, VEGF could be measured and its level was higher when cells were additionally exposed to IGF-1 and/or bFGF (figure 5). Addition of ranibizumab for 1 day did not lead to a different amount of VEGF in the medium.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Detection of VEGF in the cell culture supernatant fractions of iBREC. Presence of VEGF in the cell culture supernatant fractions of iBREC treated with bFGF IGF-1 and/or VEGF165 for 3 days was analysed by western blot. The amount of VEGF was increased slightly when cells had been treated with VEGF plus bFGF and/or IGF-1.
Discussion
In view of the accumulating evidence suggesting a crucial involvement of the growth factors VEGF, IGF-1 and bFGF in the genesis and progression of DME, we investigated their effects on the most important characteristic of REC associated with this disease: their permeability determined by the composition of TJ proteins.1–5 12 15 16 It is a distinct advantage of the cell line iBREC used in this study, that these cells—unlike primary cells—are free of contaminating cells of other types.10 Complementing rodent models of diabetes, iBREC allow accurate and detailed studies of potential higher clinical relevance as most of the involved proteins of human and bovine origin are more homologous.
In contrast to their postulated importance, IGF-1 or bFGF did not induce changes in TER and claudin-1 expression in iBREC even after long-term treatment. Like untreated iBREC, these cells when incubated with bFGF or IGF-1 did not express or secrete substantial amounts of VEGF, although weak expression of VEGF165 by primary BREC was reported.22 This difference might be due to a lower VEGF secretion of non-proliferating iBREC in confluent layers that were used in our experiments. However, a low amount of VEGF is obviously not sufficient to induce changes in TER and claudin-1 expression: whereas concentrations as low as 5 ng/ml VEGF165 stimulate proliferation and migration of iBREC, more than 20 ng/ml VEGF165 was necessary to reduce TER and claudin-1 expression (figure 1).10 15 Interestingly, combination of IGF-1 with bFGF enhanced the weak effect of VEGF165 on claudin-1 expression and TER at a moderate concentration of 25 ng/ml. This result suggests synergistic effects possible under certain conditions, which are most likely to be caused by an enhanced secretion of VEGF. The additional effect of combined action of all three growth factors seems to be relevant only in a range of VEGF165 concentrations of about 25 ng/ml in vitro, but such are the amounts measured in the vitreous of patients with diabetic retinopathy.1 3 4 In contrast to our assessment of permeability, proliferation and migration of primary BREC was strongly increased in the presence of all three factors demonstrating their more drastic synergistic effect on angiogenic processes.9
Based on our observations, we conclude that when all three growth factors are present, altered TER and TJ composition are primarily caused by action of VEGF, which can be modulated by IGF-1 and bFGF under certain conditions. This assumption is also supported by our results showing that complete reversion of decreased TER and loss of claudin-1 could be achieved by inhibition of VEGF without targeting the other factors. As expected due to the high homology of bovine and human VEGF165, ranibizumab, a VEGF-binding antibody fragment used in human therapy, efficiently reverted effects induced by long-term treatment of iBREC with VEGF165 supplemented with IGF-1 and/or bFGF, although an extended treatment was necessary (figure 4).15 16 18 23–25 Binding of ranibizumab to VEGF indeed seems to prohibit its association with the VEGF receptor(s) because the total amount of VEGF in the culture medium did not markedly change after addition of this VEGF-inhibiting agent.
It is believed that VEGF affects permeability of endothelial cells mainly by binding to KDR and its expression is differently regulated by bFGF, IGF-1 and VEGF, depending on the cells' origin and the duration of treatment.7 10 26 In our experiments, however, long-term exposure of iBREC to VEGF165 applied as single agent or in combination with IGF-1 and/or bFGF did not result in an altered expression of KDR (figures 1C and 3). Interestingly, inhibition of the tyrosine kinase activity of Flt-1 and KDR with the specific inhibitor KRN951 did not completely restore claudin-1 reduction induced by VEGF165/IGF-1/bFGF in iBREC, although the concentration used was sufficient to prevent its loss induced by VEGF165. In addition, plasma-membrane localisation of claudin-1 remained low in iBREC subsequently treated with growth factors and KRN951. Therefore, prolonged loss of claudin-1 seems not to depend exclusively on activation of KDR or Flt-1 after long-term treatment with VEGF165/IGF-1/bFGF in iBREC. Activation of the non-tyrosine kinase receptor neuropilin-1, which is also expressed in iBREC, by VEGF could lead to activation of protein kinases further downstream in the signalling pathway. However, this mechanism of bypassing KDR/Flt-1 has not been verified yet. Another explanation is based on the assumption that IGF-1 or bFGF affect permeability only at VEGF concentrations above a threshold value. Then removal of VEGF but not inhibition of VEGF receptor activity would be sufficient to reverse their effects.
In summary, our findings support the hypothesis that VEGF (but not IGF-1 or bFGF) is mainly responsible for a changed permeability in DME, but the weak synergistic effect of the three growth factors seen in our in vitro system might be important in vivo: at a VEGF concentration of low pathogenic potential, elevated concentrations of bFGF and IGF-1 might nevertheless result in destabilisation of the barrier in processes leading to DME. Even in this constellation, inhibition of VEGF was sufficient to restore their effects, which is an important result supporting preliminary data of clinical studies.24 25 In contrast to efficient targeting of VEGF, therapeutic concepts based on inhibition of tyrosine kinase activity of the VEGF receptor with a specific inhibitor seem less promising.
Acknowledgments
The authors thank Susanne Denning and Anita Ruepp for expert technical assistance and Novartis Pharma GmbH for a generous gift of ranibizumab.
References
Footnotes
Competing interests None to declare.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- At a glance