Article Text

Abstract
AIM Peripheral ulcerative keratitis (PUK) is an ocular manifestation of rheumatoid arthritis and other similar systemic diseases. The purpose of this inquiry was to investigate the involvement of matrix metalloproteinases (MMPs) in the induction and/or maintenance of PUK.
METHODS Substrate gel electrophoresis was used to characterise the MMP activities secreted by primary cultures of keratocytes derived from normal and perforated pathological corneal specimens, and those present in tears of normal subjects and patients with PUK. Substrate specificity and the in vivo activity status of the secreted MMPs was assessed by SDS-polyacrylamide gel electrophoresis of standard collagens incubated in the presence or absence of the various enzyme preparations.
RESULTS In addition to MMP-2 of M r 66 000, cultured keratocytes derived from perforated corneas of patients with PUK abnormally produce the MMP-2 of apparentM r 62 000. Other MMPs and in particular MMP-9 of M r 92 000, also occur in the tears of these patients. Their visualisation on substrate polyacrylamide gels correlated with clinical manifestations of disease activity; during periods of disease quiescence they were barely detectable. The steroid prednisolone, frequently used in systemic therapy, had no effect on the in vitro activity of MMP-2, or on its production by cultured corneal keratocytes. Although the in vitro activity of MMP-2 was inhibited by both Cu2+ and Zn2+, Cu2+ apparently induced the keratocytes to produce activated enzyme and Zn2+ irreversibly inhibited their production of MMP-2.
CONCLUSION Overexpression of corneal MMP-2 and tear film MMP-9 are characteristic features of patients with PUK and their activation may be a crucial facet of disease initiation or progression. Although effective in systemic therapy for PUK, prednisolone had no direct control over corneal MMP-2 production or activity. Zn2+ on the other hand inhibited both MMP-2 production and MMP-2 activity and may, therefore, be of therapeutic value if suitably formulated and used in conjunction with systemic steroid treatment.
- peripheral ulcerative keratitis
- keratocytes
- cornea
- gelatinase
- matrix metalloproteinases
Statistics from Altmetric.com
Peripheral ulcerative keratitis (PUK) is a chronic, progressive ocular manifestation of rheumatoid arthritis and other similar systemic diseases. Although the lesion that develops may occasionally be centrally positioned, the disorder typically progresses from a gradual thinning of the peripheral corneal stroma to the formation of a perforation prone gutter. Activated matrix metalloproteinase I (MMP-I or collagenase I), the enzyme that exhibits specificity for type I collagen, the major component of the corneal extracellular matrix, has long been implicated as a causative agent of PUK.1 2 The results of an early investigation indicated that this enzyme is secreted by limbal inflammatory cells and/or conjunctival epithelial cells lying adjacent to the corneal ulcers induced by rheumatoid arthritis.3 More recently it has been suggested that MMP-I is produced within the stroma of affected corneas, either by the corneal keratocytes themselves or by infiltrating macrophages.4 Although treatment for PUK relies heavily upon long term systemic immunosuppressive therapy,5 such treatment may not prevent corneal perforation. In this event graft surgery to replace the lysed cornea may also fail presumably because of the continued existence of activated proteases and/or inflammatory cells within the host corneal matrix. Thus, despite the general agreement that corneal destruction is inflicted by MMP-I, there exists uncertainty with respect to the source of this enzyme, its activation mechanism, and whether other proteolytic enzymes are also involved in the initiation or maintenance of the diseased state. The primary aim of the current investigation was to characterise the MMPs specifically secreted by keratocytes of perforated PUK corneas and compare their activity profiles with those secreted by keratocytes of healthy corneas. In consideration of remedial therapies for PUK, the effects of possible MMP activity regulators were also investigated. These included prednisolone, because it is currently used, but sometimes ineffectually, to arrest disease progression, and the divalent cations Cu2+ and Zn2+, because all MMPs contain Zn2+ at their catalytic centre and require Ca2+for activity. In addition to these investigations, the tears of patients suffering from rheumatoid arthritis were also assayed for MMP activity during periods of active PUK progression and periods of disease quiescence.
Methods
EXPERIMENTAL MATERIAL
Corneas
To serve as the experimental control, corneal tissue that was unsuitable for transplantation, because of minor endothelial defects, was obtained from the Bristol CTS Eye Bank. Perforated corneal tissue from patients with chronic PUK was obtained from theatre immediately after graft surgery.
Tears
After applying 20 μl of a sterile solution of saline (0.9% w/v) to the fornix of each eye, tear samples (approx 10–15 μl), were collected from healthy individuals and rheumatoid patients with PUK, using 20 μl glass capillary tubes. The clinical criteria that were used to assess the severity of the PUK in these patients are given below.
Clinical signs of active PUK progression and resolution
PUK severity relates to the degree of conjunctival inflammation, corneal thinning, and hyperaemia. The signs of active PUK are the formation of a gutter just inside the corneal limbus and considerable thinning of the cornea. While vascularisation is usually absent, there is surface staining with fluorescein dye and elevation of the conjunctiva. This often appears as a rolled edge alongside the ulcer. The signs of resolution of active inflammation include epithelialisation of the corneal surface and the formation of a superficial layer of fibrotic material beneath the epithelium. In addition, corneal vascularisation occurs, the conjunctiva flattens, and the hyperaemia resolves.
TISSUE CULTURE
Corneal keratocyte culture and maintenance
PUK corneas may contain proteolytic enzymes derived from inflammatory cells. Since the primary aim of this inquiry was to determine whether the MMPs secreted by the keratocytes of PUK corneas contributed to the destruction of their extracellular matrices, confluent keratocyte cultures were established from normal and PUK corneas as described below.
Epithelial cell layers were stripped from the underlying corneal stroma before dicing this tissue in a small quantity of Dulbecco's MEM growth medium (Gibco) supplemented with fetal calf serum (FCS, 10% v/v), glutamine (Gibco), and an antibiotic-antimycotic mixture (Sigma). The explants were seeded into 25 ml culture flasks. Once cell growth was established, to achieve uniform confluent cultures, the keratocytes were trypsinised, pelleted by centrifugation, resuspended in 4–6 ml fresh medium and divided between 4–6 culture flasks. These flasks were incubated for 1 hour at 37°C to allow the explants to adhere, then more medium (2 ml), was added. This was changed every 3–4 days. On achieving confluence, the FCS was omitted from the culture media for intermittent periods of 4 days. These media samples were then collected. Aliquots were removed and stored at −80°C for direct analysis of the secreted gelatinase activities. The protein fractions of the rest of the media were obtained by (NH4)2SO4 precipitation.
Epithelial cell culture
Ocular toxicity studies are frequently carried out using corneal epithelial cell cultures. As an addendum to the experiments in which the effects of Zn2+ and Cu2+ on keratocyte MMP secretion and viability were investigated, the effects of these metal ions on epithelial cell proliferation and viability were also assessed. The cultures were prepared by plating out corneal explants, taken from anterior limbal regions of normal corneas, in small petri dishes (5 cm diameter) and incubating them in RPM-I 1640 culture medium containing 5% v/v FCS. Once epithelial cell growth was established (∼7 days) the explants were removed to reduce the possibility of fibroblast proliferation The media were changed every 3–4 days. When near confluent, the cells were used for experimentation.
PREPARATION OF PROTEIN SAMPLES FOR MMP ANALYSIS
Keratocyte secreted proteins were harvested by (NH4)2SO4 precipitation of their serum free culture media. The (NH4)2SO4 was added as a saturated solution in 10 mM potassium phosphate buffer, pH 7.4, to a final concentration of 70% (w/v). The protein precipitates produced after a minimum period of 6 hours at 4°C were pelleted by centrifugation at 20 000 g for 30 minutes at 4°C, redissolved in 10 mM phosphate buffer, pH 7.4, containing 0.2M NaCl and 10% (v/v) glycerol and stored at −80°C (long term) or −20°C (short term).
The proteins present in tears were not concentrated. Tear solutions collected from normal subjects and patients with PUK were either assayed directly for gelatinase activity, or after storage at −80°C.
MMP DETECTION AND ACTIVITY ASSAYS
It is known that the MMPs account for over 95% of the proteolytic activity secreted by normal keratocytes.6 In vivo, these enzymes usually exist in an inactive form, either as the proenzyme, or as the activated enzyme bound to tissue inhibitor of metalloproteinase (TIMP). To cause tissue damage, they must exist for a finite period in an activated form, free of TIMP. The following methods were used to characterise, quantify, and assess the activity status of the ocular MMPs of healthy individuals and patients with PUK.
Acyl transferase activity
Since all proteases will catalyse acyl transfer independently of peptidase activity, total MMP (active and inactive) was estimated from the kinetics of liberation of 4-nitrophenol from 4-nitrophenyl acetate as described previously.6
Substrate-polyacrylamide gel electrophoresis
Gelatinase activities were separated and visualised by electrophoresis on SDS-polyacrylamide gels (8.5%) containing gelatin (1 mg/ml), as described by Unemori and Werb.7 The sample solutions contained SDS (1% w/v) and glycerol (10% v/v), and were applied to the gels without boiling or reduction. Unless otherwise stated, for characterisation of the keratocyte secreted gelatinases, equivalent volumes of the culture media of confluent cultures were applied to the gels. All other samples analysed by this method were of similar protein concentration (7 μg/lane). Protein standards of known molecular weight (M r220 000–22 000, Sigma Chem Co), were also included on each gel. After staining the gels with Coomassie blue, at applied concentrations of approximately 120 μg/ml, these proteins stained darker than the gelatin that was included in the polyacrylamide gel matrix.
Collagen hydrolysis
The in vivo state of activation of the various enzyme preparations was assessed by their ability to hydrolyse standard collagen samples. Types I, II, III, IV, and V collagens were dissolved in 10 mM acetic acid (4 mg/ml), then neutralised with an equal volume of 0.1M TRIS HCl, pH 7.8 containing 5 mM CaCl2. Denatured type I collagen (gelatin) was obtained by heating a sample of the type I collagen solution at 60°C for 20 minutes. All collagens were then incubated with and without the various enzyme preparations, in the same buffer, at 37°C. Aliquots were removed with time (0–18 hours), and added to an equal volume of the polyacrylamide gel electrophoresis (PAGE) sample buffer8 containing SDS (2% v/v) and mercaptoethanol (5%). After heating these samples at 60°C for 20 minutes, they were analysed on SDS polyacrylamide gels (10% w/v) as described by Laemmli.8
PROTEIN ESTIMATION
Protein concentration was estimated spectrophotometrically from the relation OD225 9.18 =1.0 mg/ml9 using quartz cuvettes of 1 cm path length but reduced volume. Sample protein preparations were diluted with distilled water. The solutions in which the proteins were dissolved were similarly diluted with distilled water, and used as blanks.
INHIBITION STUDIES
Effect of prednisolone, Zn2+, and Cu2+ on keratocyte MMP production and viability
For this study, the chosen experimental concentrations of prednisolone and the metal ions were effectively determined by their solubility in aqueous solution. They were dissolved in serum-free MEM culture medium at concentrations ranging from 0–1 mM and 0–2 mM respectively, before filter sterilising. The media solutions of prednisolone, Zn2+, and Cu2+ were then added, at increasing concentration, to a set of normal and a set of PUK keratocyte cultures. Each set contained keratocytes derived from the same cornea. Their culture media was harvested for MMP analysis after 4 days' incubation at 37°C. The experiment was subsequently repeated using cultures derived from different normal and PUK corneas.
Effect of Cu2+ and Zn2+ on the growth kinetics of epithelial cell cultures
Near confluent cultures of epithelial cells were trypsinised, centrifuged at 1500 rpm for 3 minutes, resuspended in the RPM-I 1640 culture medium containing 5% (v/v) FCS and counted with a haemocytometer. The cells were then pipetted into the wells of 96 well plates (50 μl aliquots of medium containing 2–5 ×103cells). After 2 hours' incubation at 37°C, a further 150 μl of this medium containing ZnCl2 (25 μm–50 mM concentration range) or CuCl2 (25 μm–2 mM concentration range) were added to each well. Each cation concentration was assayed in quadruplicate. Five replica plates were prepared and incubated at 37°C for a maximum period of 14 days. Viable cell densities were estimated at days 1, 2, 4, 7, and 14 by the hexominidase assay method of Landegren et al 10described below.
The hexominidase substrate, p-nitrophenyl-N-acetyl β-dglucosaminide, was dissolved in 0.1M citrate buffer, pH 5.0 containing 0.25% (v/v) Triton X-100 at a concentration of 7.5 mM. This was added, in 50 μl aliquots, to cells washed free of culture medium. After an incubation period of 2 hours at 37°C, the reactions were stopped with 150 mM glycine EDTA (ethylenediamine tetra-acetic acid) buffer, pH 10.4. Colour development was monitored using a microplate reader (Dynatech MR 5000) fitted with 410 nm filter. Cell densities were read against a standard curve, constructed from the results obtained from serially diluted cell suspensions of known density.
Results
CHARACTERISATION OF THE GELATINASE ACTIVITIES SECRETED BY KERATOCYTES CULTURED FROM NORMAL AND PUK CORNEAS
The gelatinase activities secreted by confluent cultures of keratocytes derived from a minimum number of 15 normal adult (45–78 years) and 10 PUK (68–82 years) corneas have been analysed by substrate polyacrylamide gel electrophoresis. Typical activity profiles of gelatinases secreted by normal and PUK keratocytes are shown in Figure 1. As expected from previously published data,6 11the major gelatinase activities detected in all samples of the keratocyte culture media could be ascribed to MMP-2. In the case of the normal keratocytes, this enzyme was represented by theM r 66 000 form predominantly. Most samples analysed using a Biorad GS-690 imaging densitometer gave readings of less than 5% for the proportion of the total gelatinase activity represented by the M r62 000 form of MMP-2 on SDS-gelatin polyacrylamide gels. (4.7 (SE 2.6); n = 7). In the case of PUK keratocytes, theM r 62 000 form of MMP-2 was much more apparent, representing between 28–45% of the total activity visualised.
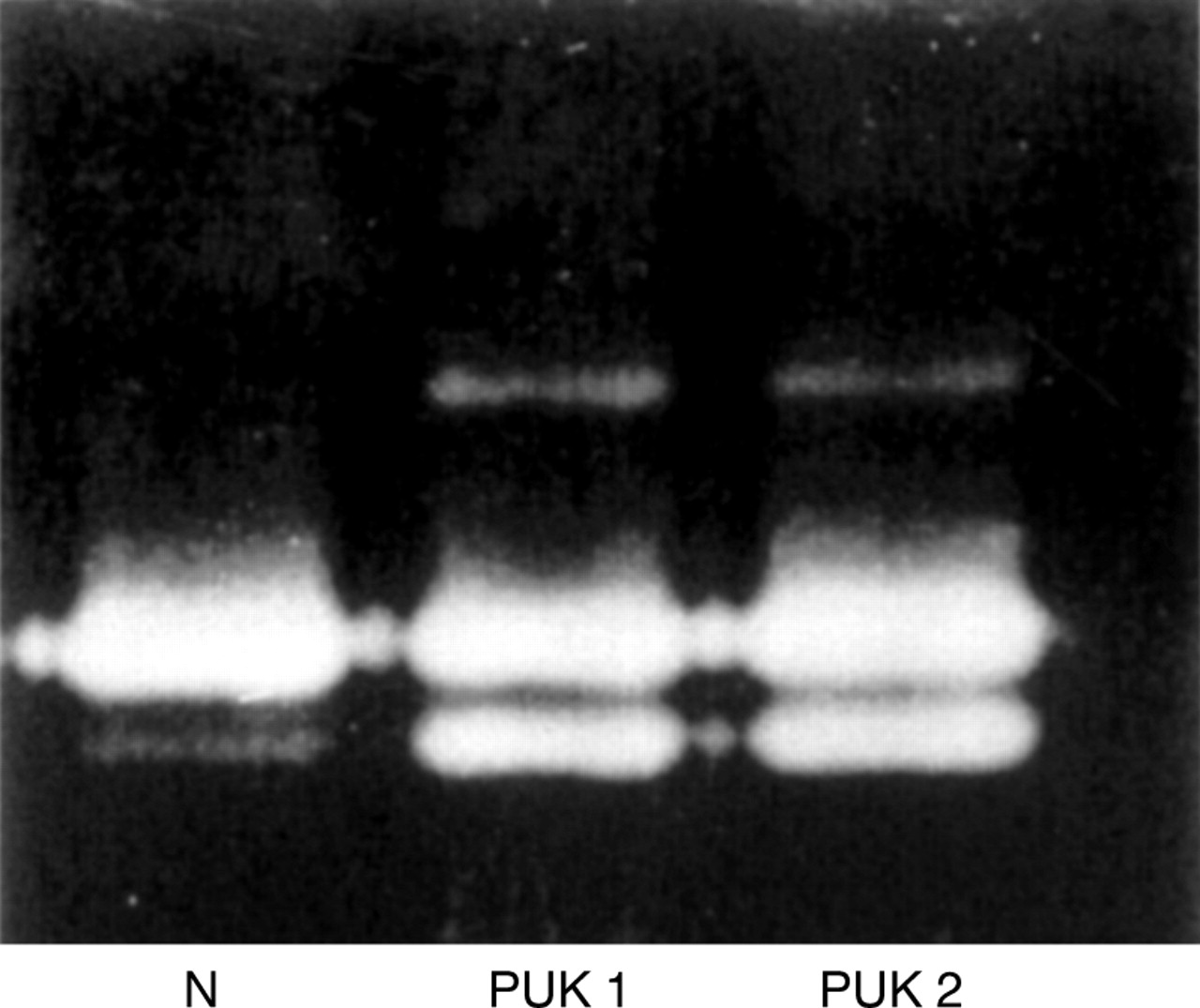
Characteristic zymographic activity profiles of the MMP-2 secreted by keratocytes cultured from normal and perforated PUK corneas.
The concentrated protein fractions of the media of several of the PUK keratocyte cultures were subsequently assayed against type I (native and denatured) and type IV collagen. Despite the obvious presence of the MMP-2 activity of M r 62 000 in the PUK keratocyte secreted protein preparations, there was no detectable degradation of any of these collagens after an incubation period of 18 hours at 37°C (data not shown).
KERATOCYTE SECRETED MMP INHIBITION STUDIES
Prednisolone
Because systemic prednisolone treatment can be an effective method of curtailing the progression of PUK, experiments were carried out to determine whether this steroid could directly affect corneal MMP activity or production.
To assess the effect of prednisolone on MMP activity itself, this steroid was added, within the limits of its solubility in aqueous solution (0–2.5 mM) to the assay buffers used to measure acyl transferase activity and develop the zymographic gelatinase activities. The data obtained by assaying the various keratocyte secreted protein preparations in the absence and presence of prednisolone are not shown, but indicated that this steroid has no effect on MMP acyl transferase activity or zymographic peptidase activity.
To assess the effects of prednisolone on MMP production, on separate occasions two different sets of normal and two different sets of PUK keratocyte cultures were incubated for 3 days in complete media then for 4 days in serum free medium containing prednisolone (0, 0.01, 0.1, and 1.0 mM) before harvesting for secreted MMP analysis. This procedure was repeated over a period of 6 weeks. During this time there was no observable change in the zymographic activities of the gelatinases secreted by either the normal or PUK cultures. Representative profiles of the latter are shown in Figure 2.
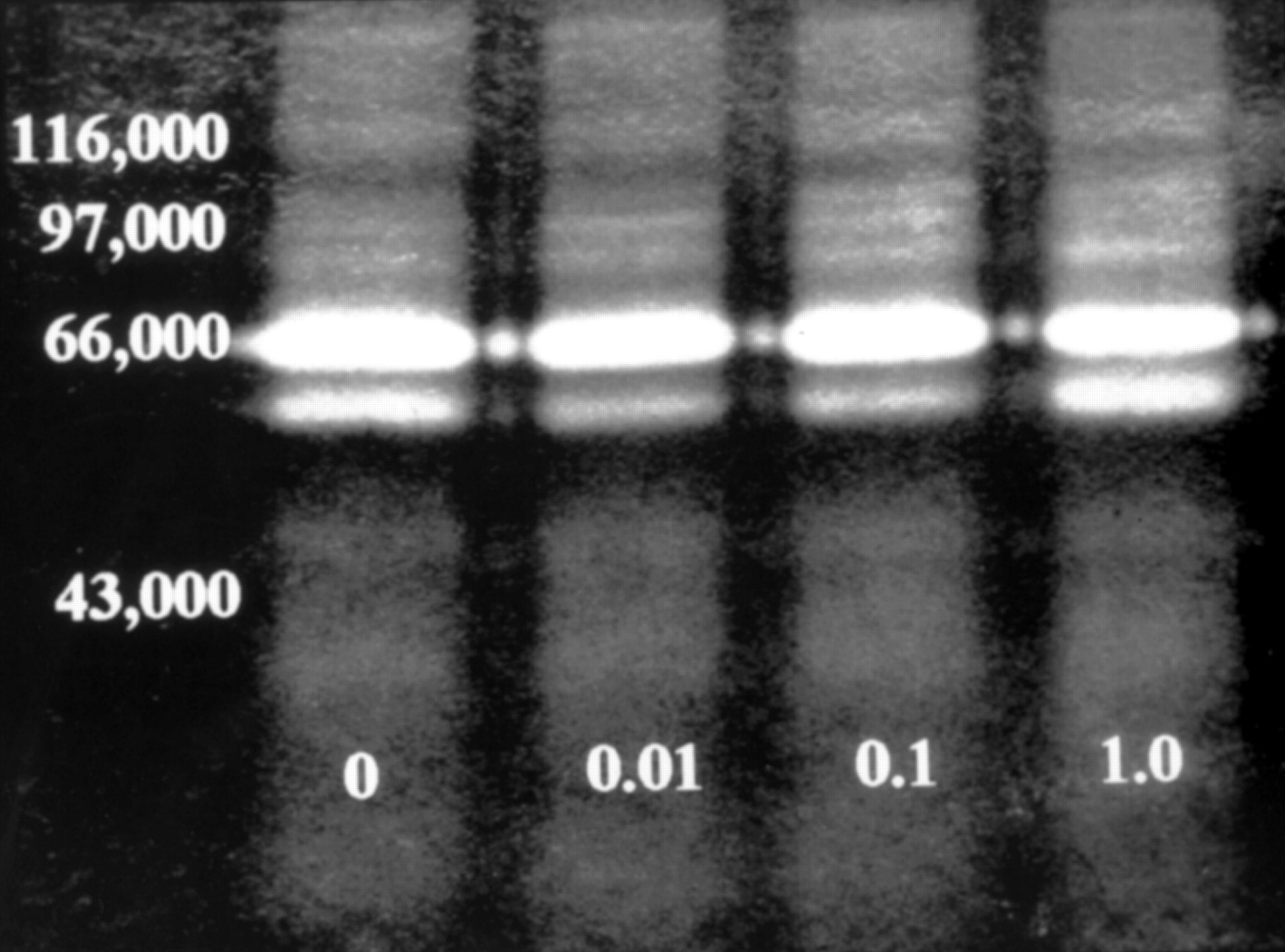
Zymographic analysis of the MMP-2 activities secreted by PUK corneal keratocytes cultured in serum free medium containing prednisolone (0, 0.01, 0.10, and 1.0 mM) for 4 days.
Cu2+ and Zn2+ cations
All MMPs contain Zn2+ at their catalytic centre and require Ca2+ for stabilising the activated form of the enzyme. Therefore, in the expectation that other divalent cations would inhibit MMP activity, the effects of Cu2+ and Zn2+ on corneal keratocyte MMP acyl transferase and peptidase activities were compared. These metal ions were included in the respective assay buffers at concentrations ranging from 0–5 mM. The kinetics of inhibition of the acyl transferase activity of the MMP secreted by normal corneal keratocytes by Cu2+ and Zn2+ are shown in Figure 3. With respect to Cu2+, the measured decrease in reaction rate was a straightforward exponential function of cation concentration (relative rate = e −0.76[Cu2+ ], ρ = 0.999). This was not the case for Zn2+; the kinetics were complex and appeared to be composed of two components. This may suggest that Zn2+ both interacts with the enzyme bound catalytic Zn2+ through the formation of zinc monohydroxide that reversibly bridges the catalytic zinc to a side chain in the enzyme's active site,12 and competes with the conformationally important Ca2+ that binds to a different domain of the protein.
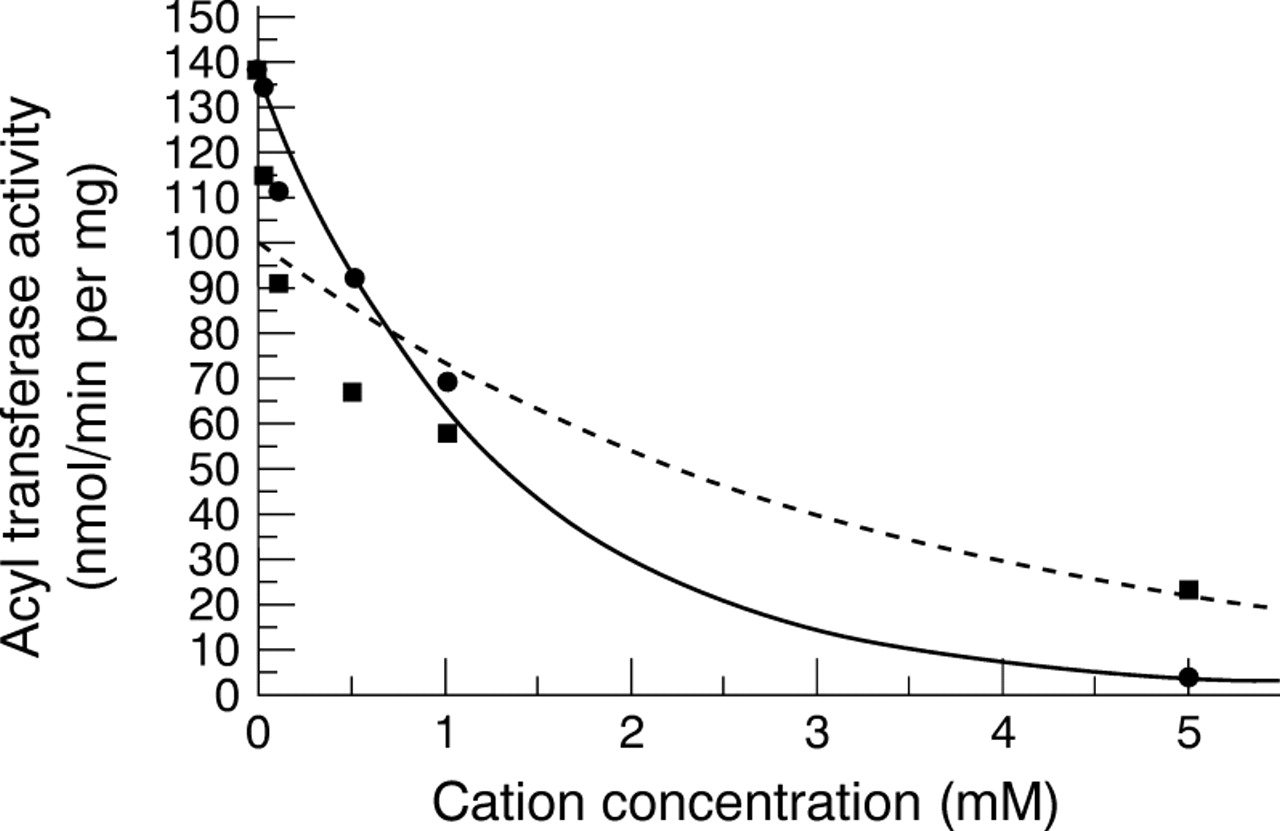
Kinetics of inhibition of acyl transferase activity of corneal MMP-2 by Cu2+ (•) and Zn2+ (•).
With respect to the effect of Cu2+ and Zn2+ on MMP peptidase activity, both metal ions completely inhibited the zymographic gelatinase activities secreted by normal and PUK corneal keratocytes at concentrations exceeding 1.0 mM. These data are not shown but, because of the observed inhibition of MMP activity in vitro, the effects of these metal ions on MMP production and on corneal keratocyte and epithelial cell viability were also investigated. They were included, at varying concentration, in the cell culture media.
The result of incubating keratocyte cultures in serum free medium in the presence of Cu2+ (1 mM) for 4 days was that they produced enzyme which, when freed of this metal ion by ultrafiltration, hydrolysed type IV but not types I, III, or V collagen (Fig 4). Analysis of these activated enzyme preparations on substrate polyacrylamide gels did not yield discrete bands of gelatinase activity: representative zymographic activity profiles of the gelatinases present in the culture media of normal keratocytes incubated with Cu2+ (0–2 mM) are shown in Figure 5A. The absence of the M r 66 000 andM r 62 000 gelatinase activities in the profiles obtained when the Cu2+ concentrations were 1 or 2 mM was very apparent and in these cases, the regions of maximum gelatinase activity occurred betweenM r 40 000 and 50 000 and at approximately M r20 000.

SDS-PAGE analysis of types I, III, IV, and V collagens incubated with aliquots of a protease preparation obtained from normal keratocyte cultures maintained in serum free medium in the presence of Cu2+ (1 mM).

Zymographic gelatinase activity profile of a normal corneal keratocyte culture maintained in serum free medium containing Cu2+ (A), and a PUK keratocyte culture maintained in serum free medium containing Zn2+ (B). In both cases the metal ion concentration ranged from 0 to 2.0 mM
The Cu2+ induced production of activated gelatinase was apparently specific to keratocytes. Whereas epithelial cell growth was unaffected by the inclusion of Cu2+ (<2 mM) in their culture media (data not shown), the physiological consequence of maintaining keratocytes in the presence of Cu2+ (>1 mM) was that they became detached from their culture flasks and eventually died (Table 1). The same phenomenon was observed when the Cu2+ induced activated gelatinase of keratocytes, prepared by ultrafiltration, was introduced into flasks containing epithelial cell cultures, but not when added to keratocyte cultures established without exposure to Cu2+.
Effect of Cu2+ on keratocyte viability and total protease secretion over an incubation period of 4 days
By contrast with the observed effects of Cu2+ on cultured keratocyte viability and gelatinase production, Zn2+ (>1 mM) inhibited MMP-2 production by keratocytes (Fig 5B) and the expected reappearance of the zymographic gelatinase activity did not occur, even after a period of 4 weeks in which the cells were maintained in the absence of Zn2+ in medium containing FCS. At the end of this period the cells appeared to be viable as judged by staining with trypan blue. Furthermore, studies in which the effect of Zn2+ on epithelial cells was assessed indicated that this cation was non-toxic to epithelial cells at the concentration that inhibited MMP-2 production. Indeed, at concentrations below 1 mM, this metal ion appeared to stimulate epithelial cell proliferation (Table2).
Effect of Zn2+ on the growth kinetics of corneal epithelial cell cultures
DETECTION AND CHARACTERISATION OF GELATINASE ACTIVITIES IN TEARS OF PATIENTS WITH PUK
For this investigation, tear samples were collected from four healthy individuals and seven patients whose severity of PUK was clinically assessed at the time of tear sample collection according to the degree of conjunctival inflammation and corneal thinning, but independently of medication. These were analysed by substrate gel electrophoresis. The data obtained indicated that normal tears contain little or no MMP but those of patients with severe symptoms of PUK all contained several high molecular weight gelatinases of apparentM r 92 000–205 000 (Fig 6). The peptidase activities of all these enzymes exhibited Ca2+ dependence and were totally inhibited by the cation chelator, EDTA at a concentration of 1 mM, and by dithiothreitol (>2 mM), even after the removal of these compounds by ultrafiltration before analysis by substrate gel electrophoresis (data not shown).
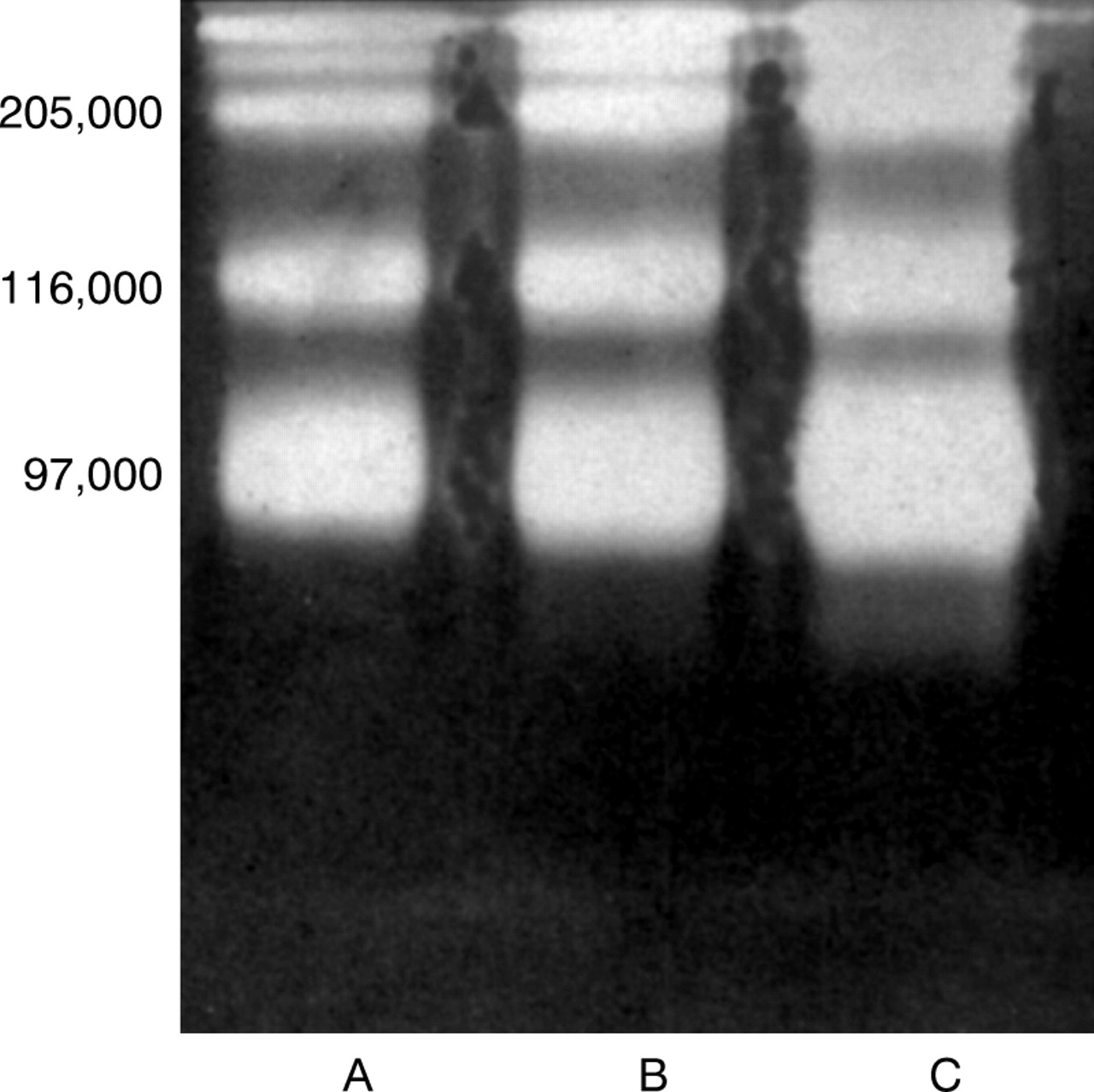
Zymographic characterisation of the gelatinase activities present in the tears of three rheumatoid patients (A, B, C) with severe PUK.
These enzymes are, therefore, MMPs. The species ofM r 92 000 co-migrated with an authentic sample of MMP-9 of M r92 000.13 The other higherM r species are also believed to be MMP-9, but complexed with TIMP (A Newby, University of Bristol, personal communication). In common with the MMPs secreted by corneal keratocytes, the MMPs in the tears of patients with PUK exhibited no detectable activity when assayed against type I, type III, type IV, and type V collagen in vitro or when assayed zymographically in the presence of Cu2+ or Zn2+ at concentrations exceeding 1 mM.
The quantity of proteolytic enzyme, including MMP-9, in tear secretions of patients with rheumatoid arthritis related to the clinical signs of advancing PUK, specifically the extent of conjunctival inflammation and corneal thinning in each eye of individual patients. The data obtained are given in Figure 7 and Table 3. From the latter, the mean (SE) values for the acyl transferase activities of proteolytic enzymes in tears of normal subjects and in tears of patients' eyes with mild, moderate, and severe PUK were calculated to be 36.5 (16) nmol/min per mg (n=8); 52 (23) nmol/min per mg(n=6); 107 (61) nmol/min per mg (n=3), and 109 (6) nmol/min per mg (n=3) respectively.

Zymographic analysis of the gelatinase activities present in the tears of rheumatoid patients with severe PUK (lane 2) and moderate PUK (lanes 1, 3, 4, 5), and in the tears of healthy subjects (lanes 6, 7).
Evaluation of the relation between clinical assessment of PUK activity and gelatinase activity in tear secretions
Statistically, the amount of proteolytic enzyme in the tears of patients' eyes with mild PUK was not significantly different from that in the tears of normal subjects (Student'st test for unpaired data, p >0.2). This was not the case, however, for the amount of proteolytic enzyme in the tears of medium and heavily inflamed eyes of PUK patients. In these tear samples, though the quantities of proteolytic enzyme were not significantly different from each other (p >0.2), they were significantly greater than that found in normal tears (p values <0.001).
INDIVIDUAL CASE STUDIES
In addition to the tear samples documented in Table 3, others were periodically collected for analysis from individual patients during the course of their treatment for PUK, in periods of exacerbation and remission. Representative zymographic activity profiles of the gelatinases present in the samples taken from one of these patients on initial presentation, during a subsequent period of quiescence brought about by systemic steroid therapy, and when the cornea finally perforated despite continuity of the systemic steroid treatment, are shown in Figure 8. From these data it was concluded that the gelatinase activity bands are most intense at the time of corneal perforation. During periods of quiescence, they become barely discernible.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zymographic analysis of the gelatinase activities in the tears of a patient with rheumatoid arthritis: (A) on initial presentation with ocular complications; (B) after a further period of 4 weeks; (C) after 4 weeks of treatment with prednisolone; (D) At time of corneal perforation.
Discussion
Peripheral ulcerative keratitis (PUK) is an ocular complication of rheumatoid arthritis. We have shown that disease progression correlates with the abnormal production of MMP-2 ofM r 62 000 in corneal stromal tissue and with the appearance of MMP-9 in tear secretions. Both these enzymes exhibit specificity for type IV basement membrane collagen.14 15 Whereas the MMP-2 is secreted by the corneal keratocytes, the MMP-9 secretory cells have not as yet been identified. It is possible that they are of the lacrimal gland, conjunctival epithelium, or the corneal epithelium itself but it is equally likely that the producer cells are the invasive inflammatory cells (macrophages and neutrophils) that accumulate at the corneal limbus.13 16 Whatever the source of MMP-9, the existence of this enzyme in tears is easily detected and quantified, and could be used as a sensitive monitor of PUK progression.
The possibility that the gelatinases (MMP-2 and MMP-9) may be involved in peripheral ulcerative keratitis has not been considered previously. Most reports concerned with the role of proteolytic enzymes in PUK have focused primarily on the stimulated production of interstitial collagenase (MMP-1) and/or an apparent reduction in the tissue concentration of its inactivating protein ligand, TIMP-1.3 4 However, because of our observation that gelatinase production or overproduction correlated with clinical manifestations of PUK progression, we hypothesised that these enzymes, once activated, may initiate perforation by breaching the corneal basement membranes (epithelial cell and Descemet's). By hydrolysing newly synthesised, non-crosslinked interstitial collagens, they could also limit tissue repair and facilitate infiltration of inflammatory cells and their proteolytic enzymes (including the MMP that hydrolyses type 1 collagen) into the corneal stroma.
If the gelatinases are involved in PUK progression, the disease may not be arrested without inhibiting their activity or production. Data presented here and elsewhere6 have indicated that keratocyte secreted protein preparations that zymographically showed the M r 66 000 andM r 62 000 activities of MMP-2 only contained no detectable gelatinase activity when assayed under native conditions. Neither was activated MMP-9 detected in tear secretions of patients with advanced disease symptoms. Although it is known that gelatinase activation involves peptide cleavage17 that may be facilitated by the action of membrane bound or MT-MMPs,18 the pathogenic mechanism of activation of these enzymes may be different. In the case of PUK, corneal perforation is a localised event and almost always occurs at a site adjacent to invasive inflammatory cells. These inflammatory cells could produce enzymes or growth factors that locally stimulate the production of activated corneal gelatinases, including the MMP-9 produced by corneal epithelial cells. Through the subsequent action of these enzymes on type IV basement membrane collagen and other susceptible components of the corneal matrix, the penetration and diffusion of macrophages and inflammatory cell proteases, including the collagenase that hydrolyses the interstitial (types I and III) collagens, would be facilitated.
In agreement with an early report3 we consider that the inflammatory cells are likely to be the source of the enzymes that cause corneal perforation. Although it has been reported that MMP-1 is an endogenous corneal enzyme, secreted by stromal keratocytes19 20 our data do not support this notion. The corneal keratocyte secreted protease preparations, which we have obtained and activated in various ways (that is, the inclusion of Cu2+ in the culture medium; protein fractionation following treatment with SDS; auto-oxidation and autocatalysis), readily hydrolyse type IV and type I collagen denatured by heating at 60°C for 20 minutes, but not native type 1 collagen.
Successful management of corneal ulcerative keratitis frequently depends upon long term treatment with systemic immunosuppressants and one of the steroids commonly employed for this is prednisolone. During periods of quiescence achieved through its administration, we have found that the gelatinases present in patients' tears also regress. However, because of the observation that prednisolone had no direct effect upon MMP-2 activity or production by corneal keratocytes, we concluded that steroid treatment may suppress the accumulation of inflammatory cells at the corneal limbus but can offer no immediate protection for corneas in imminent danger of perforating.
Although both Cu2+ and Zn2+ were shown to inhibit both the acyl transferase activity and peptidase activities of MMP-2, the effects of these cations on MMP-2 production and/or secretion by corneal keratocytes were, however, unexpectedly quite different and occurred at concentrations that were non-toxic to the growth of epithelial cell cultures (between 1 and 2 mM).
The inclusion of Cu2+ (1–2 mM) in serum free keratocyte culture media induced the secretion of activated MMPs that hydrolysed type IV and denatured type I collagen in vitro. The loss of theM r 66 000 and 62 000 forms of MMP-2 in the zymographic activity profiles of such enzyme preparations was very apparent and the regions of maximum gelatinase activity occurred between M r 40 000 and 50 000 and at approximately 20 000. These findings suggest that either MMP-2 production is inhibited by Cu2+ and that other lower M r gelatinases of similar substrate specificity are produced, or that Cu2+ induces the production of truncated but activated forms of MMP-2. The latter of these possibilities is supported by our observation that MMP-2 preparations that exhibit only theM r 66 000 and 62 000 zymographic activities never contain detectable activity when assayed under native conditions and by numerous observations, including our own, that MMP-2 activation is always accompanied by the production of a lower molecular weight gelatinase activities21-24 and in particular, one of M r 43 000. The Cu2+ induced production of activated gelatinase was apparently specific to keratocytes but the exposed cells also rounded up and detached from their matrix. Conceivably, the Cu2+also disrupted keratocyte cell junctions for, although the isolated enzyme caused cultured epithelial cells to detach from their basal matrix presumably by disrupting their basement membranes, it had no effect upon other established keratocyte cultures.
By contrast with the effect of Cu2+ on MMP production, Zn2+, within the concentration range 1–2 mM, apparently inhibited MMP production on a permanent basis. Because of this observation and the fact that Zn2+ also inhibited MMP-2 activity, it is proposed that this metal ion could be of great therapeutic value if suitably formulated for topical application.
In conclusion, the data presented in this paper indicate that ocular MMP production correlates with clinical signs of active peripheral ulcerative keratitis in patients with rheumatoid arthritis and therefore supports the possibility of their involvement in initiating and/or maintaining the disease. Because of the thermal and chemical stability of MMPs, the remedial therapy for peripheral ulcerative keratitis should include, in addition to systemic steroids, the use of specific MMP activity inhibitors or, more positively, compounds that directly inhibit MMP production and activation. Since we have shown that Zn2+ inhibits MMP-2 production by corneal keratocytes, it is suggested that this cation may be of therapeutic value for all corneal diseases that involve MMPs. Together with Cu2+ it may also be used to generate model culture systems in which the molecular events associated with MMP production and activation may be investigated.
Acknowledgments
We are very grateful to the National Eye Research Centre (NERC) for financial support.