Article Text

Statistics from Altmetric.com
Posterior capsular opacification (PCO) is the most common complication of cataract surgery, occurring in up to 50% of cases,1 and a recent clinical survey suggested that the incidence is not declining in spite of improvements in surgical technique.2 At present PCO is treated by YAG capsulotomy, which carries a small risk of sight threatening complications such as cystoid macular oedema or retinal detachment and is expensive, costing the US Medicare programme $250 million in 1993.3 YAG laser capsulotomy is frequently not available in underdeveloped countries, adding considerably to the problems of treating third world blindness from cataract. The clinical and economic significance of PCO makes it an important public health problem, and in order to prevent it a clear understanding of the pathogenesis is needed.
Cataract surgery induces a wound healing response in the lens with cellular proliferation and laying down of extracellular matrix (ECM). PCO results from the proliferation of lens epithelial cells (LEC) remaining in the capsular bag after any type of extracapsular cataract extraction.1 Visual loss occurs as a result of centripetal migration of LECs associated with fibrosis and wrinkling of the posterior capsule. Histology has shown that there are two areas within the capsular bag where cellular activity leads to the formation of PCO4—the cuboidal anterior epithelial cells and the equatorial lens bow cells, which are both derived embryologically from ectoderm at 8 weeks' gestation. Clinically there are two morphological types of PCO—wrinkling and haze from fibrosis on the capsule and epithelial pearls from regeneration of lens fibres. McDonnell and co-authors,5 6 have shown that the anterior epithelial cells in both rabbits and humans initially undergo hyperplasia and, by 4 days after surgery, transform into spindle-shaped myofibroblast cells. This process starts at the cut edge of the anterior capsule and by 1 week a ring of fibrosis forms around the rhexis where it touches the posterior capsule in the aphakic eye. The cells are surrounded by multilaminated basement membrane material composed of proteoglycans and collagen fibrils. The predominant proteoglycans found are dermatan sulphate, chondroitin sulphate, and heparan sulphate,7which are thought to determine the thickness of the collagen fibrils. The main types of collagen are I, IV, V, and VI.7
In rabbits a high proportion of the fibroblast-like cells have 4–6 nm diameter intracytoplasmic filaments which are an isoform of actin, alpha smooth muscle actin (α-SMA).8 This cytoskeleton gives these cells a retractile property and, owing to their longitudinal orientation, they may cause wrinkling of the posterior capsule which is seen in areas where cells express α-SMA. Kurosakaet al 8 found that in rabbits that had undergone cataract extraction, flattened LECs formed at the edge of the capsulorhexis by the fifth day. These cells expressed α-SMA and were related to areas of the posterior capsule where there was wrinkling.
The epithelial cells of the equator are also capable of undergoing fibrous metaplasia.1 After in the bag intraocular lens (IOL) implantation a fibrous encapsulation occurs around the haptics within the equatorial fornix. The lens bow cells of the equatorial region, however, have a stronger tendency than the anterior epithelial cells towards migration and the formation of epithelial pearls which are characterised by the presence of bladder cells.4Bladder cells appear to represent LECs that are trying to form lens fibre cells. They are 0.6–1.0 mm diameter and have a homogeneously granular cytoplasm with either a pyknotic or no nucleus. Immunohistochemistry has shown that these cells do not express α-SMA.8 In cataractogenesis, these lens bow cells can also migrate along the posterior capsule to form posterior subcapsular cataracts,9 where some of them assume the histological characteristics of bladder cells.
Cytokines involved in PCO
The histology of PCO is now well understood, but our knowledge of the cellular mechanisms that lead to these changes is incomplete. Recent research has concentrated on the cytokines that are present in the anterior segment after extracapsular cataract surgery. Cytokines are peptides secreted from cells which can then act in either a paracrine or autocrine way on their target cells. Duncanet al have studied LEC growth on human capsular bags in a protein free medium which has allowed the autocrine control by individual growth factors to be analysed.10 11Using this model they have detected basic fibroblast (b-FGF) and hepatocyte growth factors12 and transforming growth factor β (TGF-β) (I M Wormstone, personal communication). It is pertinent therefore to review the role that growth factors may have in the pathogenesis of PCO.
Transforming growth factor β
Transforming growth factor β (TGF-β) plays a central part in the cell biology of PCO. It is a serine-threonine kinase and a member of a superfamily of growth factors, including the activins and the bone morphogenic proteins, which have diverse effects on the growth and differentiation of many cell types.13 14 They are synthesised by several cell types including keratinocytes, activated macrophages, and fibroblasts and are found at particularly high concentration in platelets. The biologically active form of TGF-β is a 25 kDa dimer that is cleaved from a precursor, consisting of two disulphide linked polypeptide chains of 390 and 412 amino acids each. Cleavage of the inactive precursor by proteinases, such as plasmin, is necessary before it can bind to its receptors.13 14 The precursor binds to mannose-6-phosphate receptor which allows the plasmin to activate it.15 On release, the TGF-β dimer can be neutralised by specific binding to proteins such as α2-macroglobulin and the proteoglycans decorin and biglycan.16 Three distinct TGF-β isoforms (TGF-β1, β2, and β3) exist in humans which share a 65–85% amino acid sequence homology. The possibility that each isoform may have a biologically distinct effect is still being evaluated.
TGF-β is a multifunctional growth factor with a wide range of opposing effects on cellular processes.13 14 TGF-β inhibits epithelial cell growth but induces mitosis in mesenchymal cells such as fibroblasts. It also controls the differentiation of a variety of cell types and is important in embryogenesis, being expressed at high levels in areas undergoing rapid morphogenesis.17 It has been shown to induce cataractous changes in rat lenses.18 In corneal epithelium, the expression of TGF-β and the receptors for β1 and β2 have been shown to be upregulated during wound healing following excimer laser ablation.19 TGF-β is also important in modulating the production and remodelling of extracellular matrix and it also controls cell-cell and cell-matrix interactions. The regulation of cell growth, matrix production, and the inflammatory response suggests that TGF-β has a central role in the wound healing process.
Source of TGF-β
Pasquale et al 20investigated the distribution of TGF-β in the anterior segment of the human eye by immunohistochemistry. TGF-β2 was more widely distributed than TGF-β1 and TGF-β3 was not found in ocular tissues. Both TGF-β1 and β2 were found in the proximal portion of the ciliary processes and the limbal epithelium, where it has been postulated to be involved in the transdifferentiation of conjunctival to corneal epithelium. TGF-β2 is probably produced locally since the predominant isoform in serum is TGF-β1. Human LECs produce TGF β21in vitro, and both TGF-β1 and β2 have been immunolocalised to equatorial and anterior LECs, with relatively stronger labelling at the equator.22 The equatorial cells also expressed the receptors for β1 and β2, indicating that there is autocrine stimulation. TGF-β has been assayed in human and rabbit aqueous humour. Cousins et al found the total concentration of human TGF-β to be 450 pg/ml.23 Ninety per cent was TGF-β2 which had a fourfold greater activity on inhibition of epithelial cell mitosis than TGF-β1. Jampelet al 24 found the amount of human aqueous TGF-β2 ranged from 2.3 to 8.1 ng/ml, with 60% being in the active form. The concentration of active TGF-β has been measured in rabbit aqueous during the first month after cataract surgery.25 There was a sevenfold decrease in concentration on day 1 from preoperative values which might have been due in part to increased aqueous protein from breakdown of the blood-aqueous barrier. A concomitant increase in aqueous α2-macroglobulin was likely to neutralise the action of TGF-β. The concentration of active TGF-β returned to normal preoperative concentration on day 15 and remained so for 1 month after surgery.
Cell proliferation
Cellular proliferation is controlled by the action of both stimulatory and inhibitory molecules. TGF-β reversibly inhibits cell division by acting negatively on the cell cycle by arresting progression through the late portion of the G1 phase.26 27 Early studies showed that TGF-β2 inhibited LEC proliferation in vitro (Table 1) TGF-β2 from human aqueous humour had an inhibitory effect on bovine LEC proliferation, and the addition of anti-TGF-β2 monoclonal antibody abolished this effect.28 Using rabbits that had undergone cataract surgery, the effect of preoperative and postoperative aqueous humour on the proliferation of rabbit LECs in vitro was studied.25Aqueous humour obtained from rabbits that had undergone surgery had a significantly greater proliferative effect on LEC growth compared with preoperative aqueous. The effect was greatest on day 1, decreasing with time postoperatively as the concentration of active TGF-β increased. It was concluded that an initial fall in TGF-β concentration allowed the proliferative action of other growth factors, such as basic fibroblast growth factor (b-FGF) and epidermal growth factor (EGF) to occur (see Fig 2). Once active TGF-β had returned to preoperative levels (by 2 weeks) it was postulated that the proliferative effect of the other cytokines in the aqueous was suppressed.
Cytokines involved in the pathogenesis of posterior capsule opacification

Concentration and actions of TGF-β and b-FGF following cataract surgery in rabbits.
Cell differentiation
A notable feature of the fibrotic form of PCO is that it can cause wrinkling of the capsule as well as capsular contraction which can lead to intraocular lens decentration and visual disturbance.1It has been established that this effect is caused by LECs, in particular the anterior LECs, which express α-SMA. Numerous studies have looked at the effect that growth factors have on the expression of α-SMA in different types of cells. For example, the subcutaneous administration of TGF-β1 in rats caused the formation of granulation tissue, in which myofibroblasts expressing α-SMA were abundant.29 Neutralising antibodies to TGF-β1 abolished the increased expression of α-SMA by TGF-β1 (Table 1) Kurosakaet al showed that culturing bovine LECs on a collagen gel caused its shrinkage through shortening of the collagen fibrils30 and showed that TGF-β2 increased both collagen gel contraction, in a dose dependent manner, and the ratio of α-SMA expressing LECs. The formation of myofibroblasts 4 days after surgery8 suggests that TGF-β acts within the first week postoperatively, even though it has been shown that active TGF-β concentration is low during this time.25 Alternatively, an unknown cytokine may stimulate myofibroblast formation early in the postoperative period.
Extracellular matrix
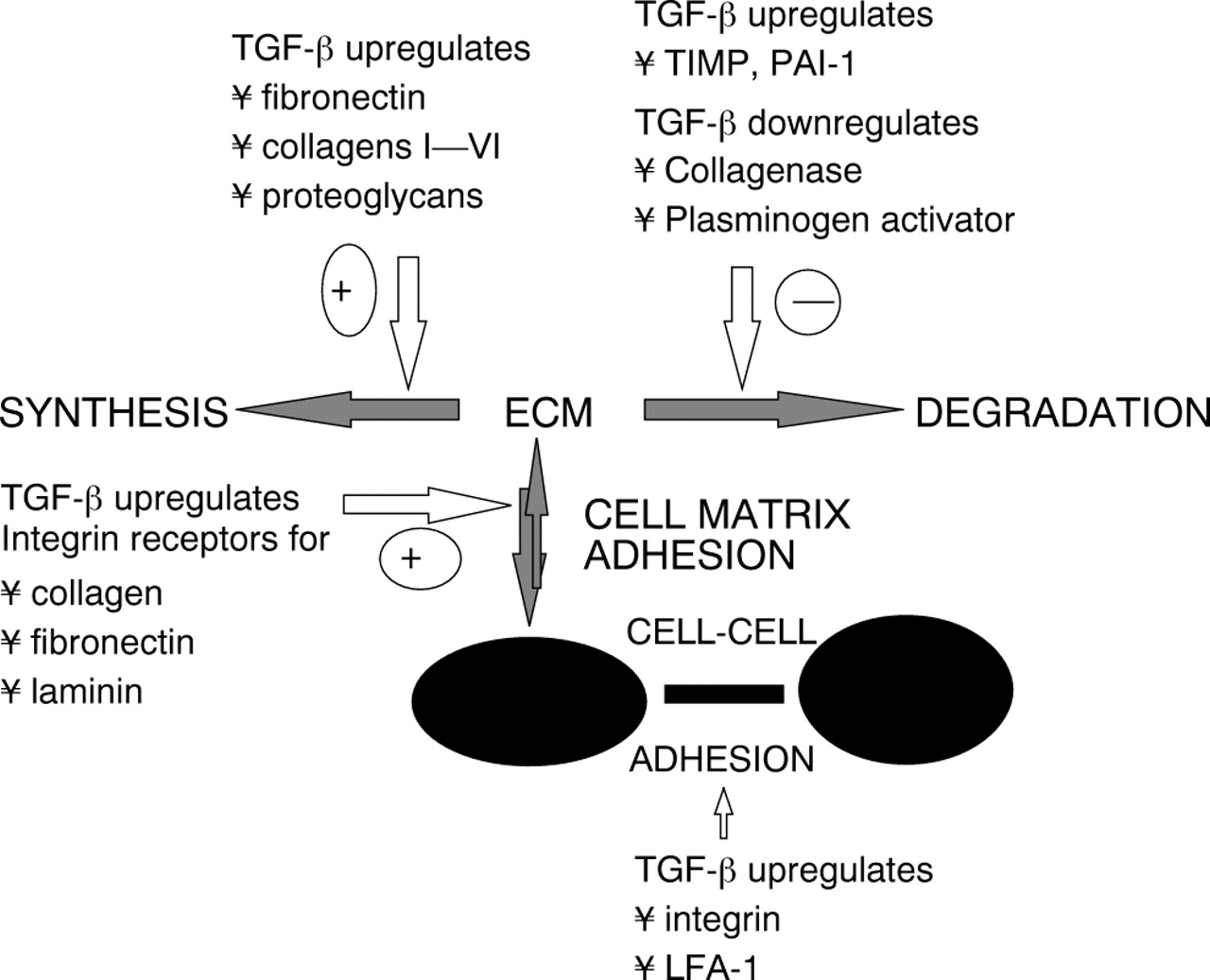
An important aspect of the wound healing response is the laying down of extracellular matrix (ECM) by LECs. TGF-β has been shown to regulate the expression of certain ECM components including proteoglycans, fibronectin, and collagen, as well as the cell surface receptors that bind the matrix proteins31 (Table 1). At picomolar concentrations, TGF-β increased the expression of chondroitin and dermatan sulphate in fibroblasts up to 20-fold. It increased the biosynthetic rate of the 45 kDa proteoglycan core protein and the molecular mass of the glycosaminoglycan chains in both molecules. In PCO, the collagen within the ECM is secreted by the LECs (both anterior and equatorial) and TGF-β increases this.32
The turnover of the ECM is determined by the balance between the ECM degrading enzymes, matrix metalloproteinases (MMPs), and the tissue inhibitors of matrix metalloproteinases (TIMPs) which suppress them (Fig 1). The major MMPs include: MMP-1 (collagenase), MMP-2 (gelatinase A), MMP-3 (stromelysin-1) and MMP-9 (gelatinase B).13MMP-2 is involved in the degradation of collagen IV. TGF-β has been shown to decrease the expression of collagenase and to increase the collagenase inhibitor TIMP in human fibroblasts, thereby augmenting the formation of ECM.33 In addition, TGF-β increased the levels of plasminogen activator inhibitor type 1, thereby indirectly reducing collagenolytic activity by inhibiting the activation of plasminogen to plasmin, an activator of procollagenase.

{kind=link}
{kind=link}
Role of TGF-β in the turnover of extracellular matrix.
Cell adhesion
LEC hyperplasia is dependent on firm attachment to components of the posterior capsule. Cell adhesion is mediated by a set of cell surface receptors called integrins, which are also regulated by TGF-β1.34 The integrins are a family of membrane spanning glycoproteins that interact with cellular actin and thus link the ECM to the cellular cytoskeleton. Their basic structure is a heterodimeric complex of one 130–200 kDa α subunit linked non-covalently to one 90–130 kDa β subunit.35 Binding of the extracellular proteins requires the presence of Ca2+or Mg2+ within the α chain while the β subunit recognises an arginine-glycine-aspartic acid sequence in the ECM protein. Human LECs express β1 integrin and other cell adhesion molecules (CAMs)—I-CAM 1 (intercellular adhesion molecule 1) and CD44.36 Growth in vitro on collagen IV/laminin coated plates was inhibited by addition of monoclonal antibodies to CAMs indicating that LECs use CAMs to attach to laminin and collagen IV. Since laminin and collagen IV are major components of the lens capsule which promote LEC adhesion37 it is likely in vivo that CAMs also serve in the attachment of LECs to the capsule.
Recently the sustained delivery of the RGD peptide (arginine-glycine-aspartic acid sequence) has been shown to reduce PCO38 by competitively inhibiting the binding of the LECs to the ECM via the β1 integrin present on the LEC surface. The sustained release of RGD peptide in the capsular bag in rabbits following cataract surgery has been shown to reduce the migration of LECs on the posterior capsule.38 It remains to be confirmed whether this is also the case with human LECs.
Apoptosis
Liu et al 39 have shown that rat LECs cultured in the presence of TGF-β underwent apoptosis. The number of myofibroblast-like cells decreased within days of their initial proliferation by apoptotic cell death, defined by electron microscopy, and it was hypothesised that TGF-β might be important in this process. Kato and co-workers looked at LECs in rabbits following lens extraction40 and found that apoptosis occurred only in the LECs that differentiated into myofibroblasts at the incised edge of the anterior capsule. They suggested that because TGF-β plays such an important part in myofibroblast differentiation, the same cytokine causes apoptosis. The assay they used to detect apoptotic cells (TUNEL assay which detects fragmented DNA) does not distinguish these from necrotic cells, since DNA fragments form in both. This can be done by measuring the integrity of the plasma membrane, which is intact in cells undergoing apoptosis but breached in necrosis. It may be that necrosis is more important in stimulating wound repair since this will result in the release of cellular cytokines from damaged cells which does not occur in programmed cell death. It is therefore important to distinguish both forms of cell death. If apoptosis is an important factor in the pathogenesis of PCO, then protocols might be developed to prevent re-epithelialisation through this process.
Other cytokines
BASIC FIBROBLAST GROWTH FACTOR (B-FGF)
Basic fibroblast growth factor (b-FGF) is a polypeptide which influences the proliferation and differentiation of a variety of cells derived from neuroectoderm, mesoderm, and neural crest. It binds heparin which is necessary for recruitment to its receptor on the cell surface and action. It is particularly potent in the control of fibroblast cell growth and angiogenesis.41-43 It is present in the lens and is involved in its development.44In ectodermal and mesodermal cells it regulates cell proliferation and migration. It stimulates lens fibre differentiation in a dose dependent manner.45 46
b-FGF has a molecular weight of 17 kDa47 at a concentration of 1.074 (SD 0.158) mg/ml in human aqueous.48 Hayashi and Kato have demonstrated that human LECs express b-FGF in vitro.49 Wallentinet al showed that aqueous b-FGF was increased in rabbits up to 30 days following cataract extraction.25 b-FGF was found to stimulate LEC mitosis and increase the formation of collagen by these cells in culture.32 TGF-β2 has been shown to inhibit the proliferative effect of b-FGF on rabbit LEC growth.25b-FGF reduced the contraction of a collagen gel by bovine LECs and the proportion of cells expressing α-SMA,30 indicating that b-FGF has opposing actions to TGF-β2 (Table 1). However, in this study no interaction between b-FGF and TGF-β2 was investigated.
INTERLEUKIN-1 AND 6
Interleukin-1 (IL-1) is a mediator of intercellular signals within the immune system and between the immune system and other organs.50 The predominant form is a 17 kDa glycoprotein which is produced by a variety of cell types including monocytes, macrophages, lymphocytes, and fibroblasts. It activates various cells which then release a cascade of inflammatory mediators51and has multiple effects on cells involved in inflammation and wound healing, even at concentrations of 10−10–10−12 M.
IL-1 is synthesised by human LECs in vitro52 and stimulates LEC mitosis and collagen synthesis by these cells.32 Both of these effects were reduced by a monoclonal antibody to the IL-1 receptor. IL-1 increases the synthesis of prostaglandin E2 (PGE2) by cells, through the activation of phospholipase A2 and cyclo-oxygenase in the metabolism of arachidonic acid53 and stimulates human LECs to produce PGE2 which contributes to increased inflammation after cataract surgery. Using human LECs in vitro, the concentration of PGE2 in supernatants increased between 1 week and 1 month as the LECs proliferated and underwent fibroblastic changes.52 The cyclo-oxygenase inhibitor, diclofenac, decreased PGE2 synthesis from human LECs in vitro54 and at the same time inhibited LEC mitosis and collagen synthesis. However, this may not be an important mechanism as animal studies have shown that the sustained delivery of diclofenac following cataract surgery failed to prevent PCO.55
The interleukin, IL-6, is produced, as a 26 kDa polypeptide, by monocytes, epithelial cells, and fibroblasts. The major effects of IL-6 described are the proliferation and differentiation of cells as well as increasing secretion of acute phase proteins by the liver.56 It induced uveitis in rabbits when injected intracamerally.57 In the same report, 3 days after routine cataract surgery in 12 patients, the aqueous concentration of IL-6 was as high as 3 μg/ml, a 4000-fold increase on preoperative levels. IL-1 induced human vein endothelial cells to secrete IL-658suggesting that the observed actions of IL-1, already described, may be mediated through the production of IL-6. However, there has as yet been no study of IL-6 action on LECs.
Conclusion
Even though most of the information on growth factors comes from animal studies, it currently appears that TGF-β plays an influential part in the genesis of PCO (Table 1) in humans. Immediately after surgery, the low levels of active TGF-β may allow b-FGF to stimulate LEC proliferation (Fig 2, Table 1). When the active TGF-β returns to normal levels, at around 2 weeks after surgery, it serves as a stop signal for proliferation but, simultaneously, as a stimulus for the other characteristic PCO changes, such as myofibroblastic differentiation and ECM formation, and attachment of LECs to the underlying posterior capsule. Histological studies have shown, however, that these events occur within the first week postoperatively, indicating that other cytokines must be involved.
Regulation of TGF-β may prove an effective means of inhibiting PCO. Following cataract extraction, LECs are exposed to active TGF-β and the administration of a biological agent or drug to either antagonise its effect or block its action could be useful therapeutically. This therapeutic effect would be needed while active TGF-β is present (for up to 1 month), but the molecular interactions between it and other growth factors, such as b-FGF on LEC mitosis, must be considered.
As with TGF-β, the biological effects of b-FGF during the production of PCO may be both detrimental or beneficial, since it stimulates LEC mitosis and collagen synthesis, but reduces the expression of α-SMA. The ratio of both growth factors may be important in influencing LEC behaviour after cataract surgery.
Since cataract surgery causes a transient increase in inflammation postoperatively, the triggering effects of the inflammatory mediators, such as IL-1 and IL-6, may have an early stimulating effect on PCO formation. However, there has been no conclusive evidence that an anti-inflammatory drug, such as diclofenac, will be of benefit in preventing PCO.
Finally, one has to remember that growth factors can have different effects in animals59 than in humans and that much more research using human models is needed.
Acknowledgments
The authors wish to thank Michael Wormstone for his advice and help.