Article Text

Abstract
Background/aims Pseudoexfoliation syndrome (PEX) is characterised by the production and accumulation of extracellular fibrillar material in the anterior segment of the eye. The pathogenesis of PEX is multifactorial with genetic factors and ageing as contributing factors. Previously, an increased concentration of beta-crystalline B2 (CRYBB2) was observed in the aqueous humour (AH) in eyes with PEX in a pooled material. Here, the protein content was examined on individual basis.
Methods During cataract surgery, AH was sampled from patients with and without PEX, 10 eyes in each group. The proteins were digested and labelled with isotopomeric dimethyl labels, separated with high-pressure liquid chromatography and analysed in an Orbitrap mass analyzer.
Results The concentration of complement factor 3, kininogen-1, antithrombin III and vitamin D-binding protein was increased in all eyes with PEX. Retinol-binding protein 3, glutathione peroxidase, calsyntenin-1 and carboxypeptidase E were decreased in eyes with PEX. Beta-crystalline B1 and CRYBB2 and gamma-crystalline D were up to eightfold upregulated in 4 of 10 in eyes with PEX.
Conclusion The results indicate that oxidative stress and inflammation are contributing factors in the formation of PEX. Knowledge about the proteome in PEX is relevant for understanding this condition.
- aqueous humour
- diagnostic tests/Investigation
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
The pseudoexfoliation syndrome (PEX) is an age-related disorder characterised by the production and accumulation of extracellular fibrillar material in the anterior segment of the eye. Patients with PEX have an increased risk of developing glaucoma and increased risks for complications during and after cataract surgery in patients.1 2 The pathogenesis of PEX is not yet fully understood, but it is presumably multifactorial with ageing and genetics as contributing factors.1 3 Several investigators have provided evidence that oxidative stress is involved in the pathogenesis of PEX. Decreased concentrations of antioxidants and increased concentrations of oxidative stress markers have been described in the aqueous humour (AH) in eyes with PEX.4 In a previous study, the protein content of AH in eyes with PEX compared with controls was investigated using isobaric tags for relative and absolute quantification, proceeded by capillary liquid chromatography and capillary electrophoresis, followed by matrix-assisted laser desorption/ionisation-time of flight tandem mass spectrometry (MS/MS). In the pooled AH material, an increased concentration of beta-crystalline B2 (CRYBB2) was observed in eyes with PEX compared with controls.5 The present study is an extension of our previous studies, expanding the knowledge about PEX on an individual level. Isotopomeric dimethyl labelling of samples permits quantitative studies on the protein content on an individual basis. The aim of the present study was to investigate and quantify the protein content in AH in individuals with PEX and in eyes with cataract only.
AH was sampled during cataract surgery from patients with and without PEX. The proteins were digested according to the filter-aided sample preparation protocol (FASP) and the peptides labelled with isotopomeric dimethyl labels.6 7 The samples were separated using high-pressure liquid chromatography and analysed further in an Orbitrap mass spectrometer.
Materials and methods
Patients
The samples consisted of AH from 11 eyes with PEX syndrome (11 patients). They had no other ophthalmic disease except cataract and PEX. Samples from 11 eyes (11 patients) with cataract only were used as controls. All patients underwent a comprehensive ophthalmic examination prior to cataract surgery. All eyes with PEX had normal intraocular pressure defined as <21 mm Hg, and no ophthalmic condition other than cataract. None of the patients were on ophthalmic medication in either eye. Patients with previous laser or intraocular surgery, or systemic conditions, such as diabetes, were not included in the study. The age-matched control group had no other ophthalmic condition other than cataract and fulfilled all the criteria described above. Informed consent was obtained from patients prior to collecting the samples and the study was approved by the Ethics Committee of Uppsala University (2005:109). The study followed the tenets of the Declaration of Helsinki.
Sample collection
From the anterior chamber, a small quantity of AH, 0.10 mL was withdrawn through a paracentesis in clear cornea with a 27-gauge needle on a tuberculin syringe (Oasis, Glendora, CA, USA). The samples were immediately transferred to sterile plastic tubes (Thermowell tubes 6571, Coster, England) and stored at –80°C until analysis.
Protein sample preparation and mass spectrometry
Samples were freeze-dried over night and dissolved in sodium dodecyl sulphate (SDS) lysis buffer (4% SDS in 100 mM Tris/HCl, pH 7.6) at 70°C for 5 min. Solubilised proteins were separated from sample debris by centrifugation at 16 000 g for 5 min, and protein concentration was determined by detergent compatible protein assay (Bio-Rad, Hercules, CA, USA). Enzymatic fragmentation of proteins was performed by FASP.6 In brief, protein disulphide bonds were reduced with 100 mM dithiothreitol at 37°C for 30 min prior to sample loading onto a 30 kDa centrifugal filter unit (Millipore, Merck, Germany). Next, samples were washed with urea buffer (8 M urea in 100 mM Tris/HCl, pH 8.5) and alkylated with 550 mM iodoacetamide. Protein digestion was performed by Lys-C (protein-to-enzyme ratio 100:1; Wako Chemicals GmbH, Neuss, Japan) at 30°C for 2 hours, followed by trypsin (protein-to-enzyme ratio 100:1; Promega Corporation, Fitchburg, WI, USA) at 37°C over night. Resulting peptides were eluted with 50 mM tetraethylammonium bromide, acidified and dried before further processing.
To perform a quantitative proteome analysis, peptides were modified by stable isotope dimethyl labelling according to Boersema et al.7 In brief, control samples were light labelled in 4% formaldehyde and 0.6 M sodium cyanoborohydride for 1 hour at room temperature. For heavy labelling of PEX samples, stable isotope substituted formaldehyde (CD2O) was used. Next, labelling reaction was stopped by acidification with 1% ammonia solution and 5% formic acid, prior to mixing heavy (PEX) and light (control) samples. For sample clean-up and purification before MS analysis, SPE Isolute C18 columns (Biotage, Uppsala, Sweden) were used.
Reverse-phase chromatography for peptide separation was performed using an Easy nano flow system (Thermo Fisher Scientific) coupled to an LTQ-Orbitrap Velos Pro mass spectrometer (Thermo Fisher Scientific). Peptides were separated by precolumn (100 µm ID, 5 µm C18 beads) and analytical columns (75 µm ID, 3 µm C18 beads) (Thermo Fisher Scientific), using a linear gradient from 4% to 48% acetonitrile with 0.1% formic acid for 221 min at a flow rate of 250 nL/min, followed by 75% acetonitrile for 10 min and 4% acetonitrile for 9 min for re-equilibration. After separation, peptides were ionised using a nano-electrospray ionisation source and transferred into the mass spectrometer. Full survey scan spectra (m/z=400–1750) were acquired in the Orbitrap with a resolution of 60 000 after accumulation of 1 000 000 ions. The 10 most intense peaks were isolated and fragmented in the linear ion trap using collision-induced dissociation (35% normalised collision energy). Mass spectrometer worked in data-dependent mode.
Raw data were processed using MaxQuant. Database searches were performed using the implemented Andromeda search engine to correlate MS/MS spectra to the updated Uniprot human database. The following parameters were used for data processing: maximum of two miss cleavages, mass tolerance of 4.5 ppm for main search, trypsin as digesting enzyme, carbamidomethylation of cysteins as fixed modification, oxidation of methionine and acetylation of the protein N-terminus as variable modifications. For dimethyl labelling, Lys0 and Nter0 were set for light label, and Lys4 and Nter4 for heavy label. Only peptides with a minimum of seven amino acids as well as at least one unique peptide were required for protein identification. For quantification of proteins, two ratio counts were set as a minimum. Only proteins with at least two peptides and at least one unique peptide were considered as identified and used for further data analysis. Perseus software (Max Planck Institute of Biochemistry, Martinsreid, Germany; http://www.perseus-framework.org) was used for statistical testing and p values were corrected for multiple testing using Benjamini-Hochberg method.
Results
For individual proteome characterisation of AH, samples of 11 patients with PEX syndrome and 11 patients with cataract only as controls were collected and processed. One experiment gave overall very low ion signals and was therefore excluded in further analysis. Thus, 10 PEX–control matched pairs remained for further analysis. They consisted of samples from 10 eyes with PEX syndrome (10 patients), seven females and three males with a mean age of 80 years (range 65–84). The controls consisted of AH from 10 eyes (10 patients), seven females and three males with a mean age of 73 years (range 59–85). Proteins were extracted in SDS lysis buffer and digested according to FASP with Lys-C and trypsin. To achieve a quantitative proteome analysis, peptides were labelled using stable isotope dimethyl labelling, mixed in a one-to-one ratio, PEX as heavy and control as light labelled samples, and measured using a high-resolution mass spectrometer (LTQ-Orbitrap Velos Pro). In average, we identified 110 proteins in each sample (99–119 proteins per sample), which lead to a total identification of 209 proteins out of 10 replicates of AH (figure 1). Out of these, 184 proteins were quantified based on heavy-to-light ratios.

Cumulative number of identified proteins out of 10 individual patient samples. AH of 10 patients with PEX and 10 control patients with cataract only were analysed by high-resolution mass spectrometry including quantification based on stable isotope dimethyl labelling. In total, 209 nonredundant proteins were identified. AH, aqueous humour; PEX, pseudoexfoliation syndrome.
To characterise the proteome of AH of patients with PEX syndrome, quantitative proteomics based on stable isotope dimethyl labelling was performed. For statistical evaluation, we considered proteins which were quantified in at least 8 out of 10 replicates. Hence, a total number of 71 proteins were quantified in patients with PEX and control samples as illustrated in figure 2. The group of proteins, which were significantly upregulated in samples of PEX patients, consisted of complement C3 (C3; fold change, FC: 0.3), kininogen-1 (KNG1; FC: 0.2), antithrombin-III (SERPINC1; FC: 0.2) and vitamin D-binding protein (GC; FC: 0.1). Significantly downregulated proteins included calsyntenin-1 (CLSTN1; log2 FC: −0.8), retinol-binding protein 3 (RBP3; FC: −0.9), carboxypeptidase E (CPE; FC: −0.9) and glutathione peroxidase 3 (GPX3; FC: −0.8) (figure 2).
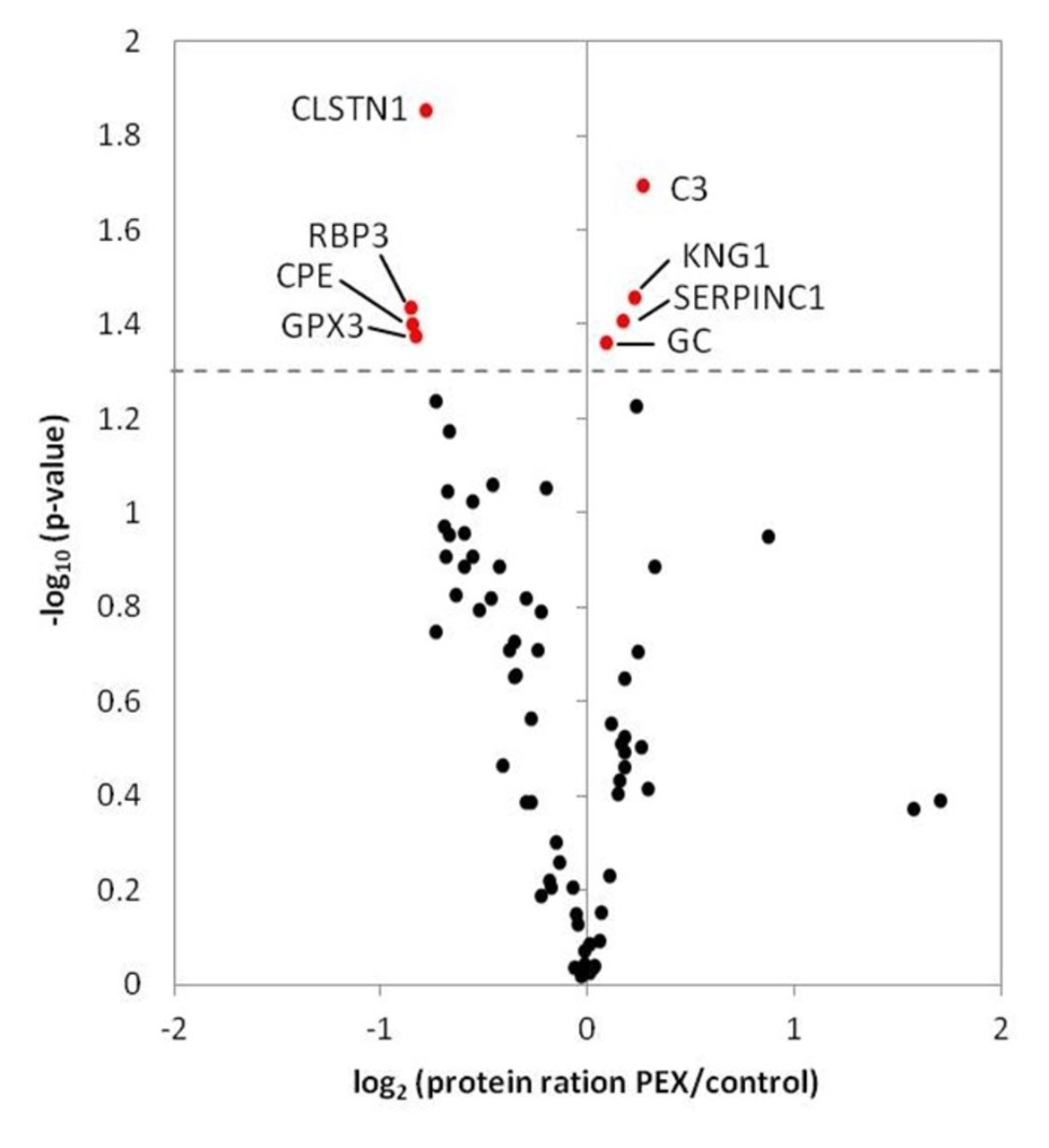
Protein quantification of PEX and control samples based on stable isotope dimethyl labelling.Volcano plot includes 71 proteins which were quantified in at least 8 out of 10 replicates. Protein rations (log2) were plotted against negative logarithmised p values (−log10). Significantly regulated proteins (p<0.05) are coloured in red. Significantly upregulated proteins included C3, KNG1, antithrombin-III (SERPINC1) and vitamin D-binding protein (GC). Significantly downregulated proteins included CLSTN1, RBP3, CPE and GPX3. C3, complement C3; CLSTN1, calsyntenin-1; CPE, carboxypeptidase E; GPX3, glutathione peroxidase 3; KNG1, kininogen-1; PEX, pseudoexfoliation syndrome; RBP3, retinol-binding protein 3.
Next, we analysed our data by considering protein quantification in at least 4 out of 10 replicates and identified a group of three significantly and strongly upregulated proteins (figure 3). All these proteins were crystallins; beta-crystalline B1 (CRYBB1; FC: 8.1), CRYBB2 (FC: 6.0) and gamma-crystalline D (CRYGD; FC: 3.9). To illustrate the strong regulation, protein intensities of control samples were plotted against protein intensities of PEX samples of each crystalline (figure 4).
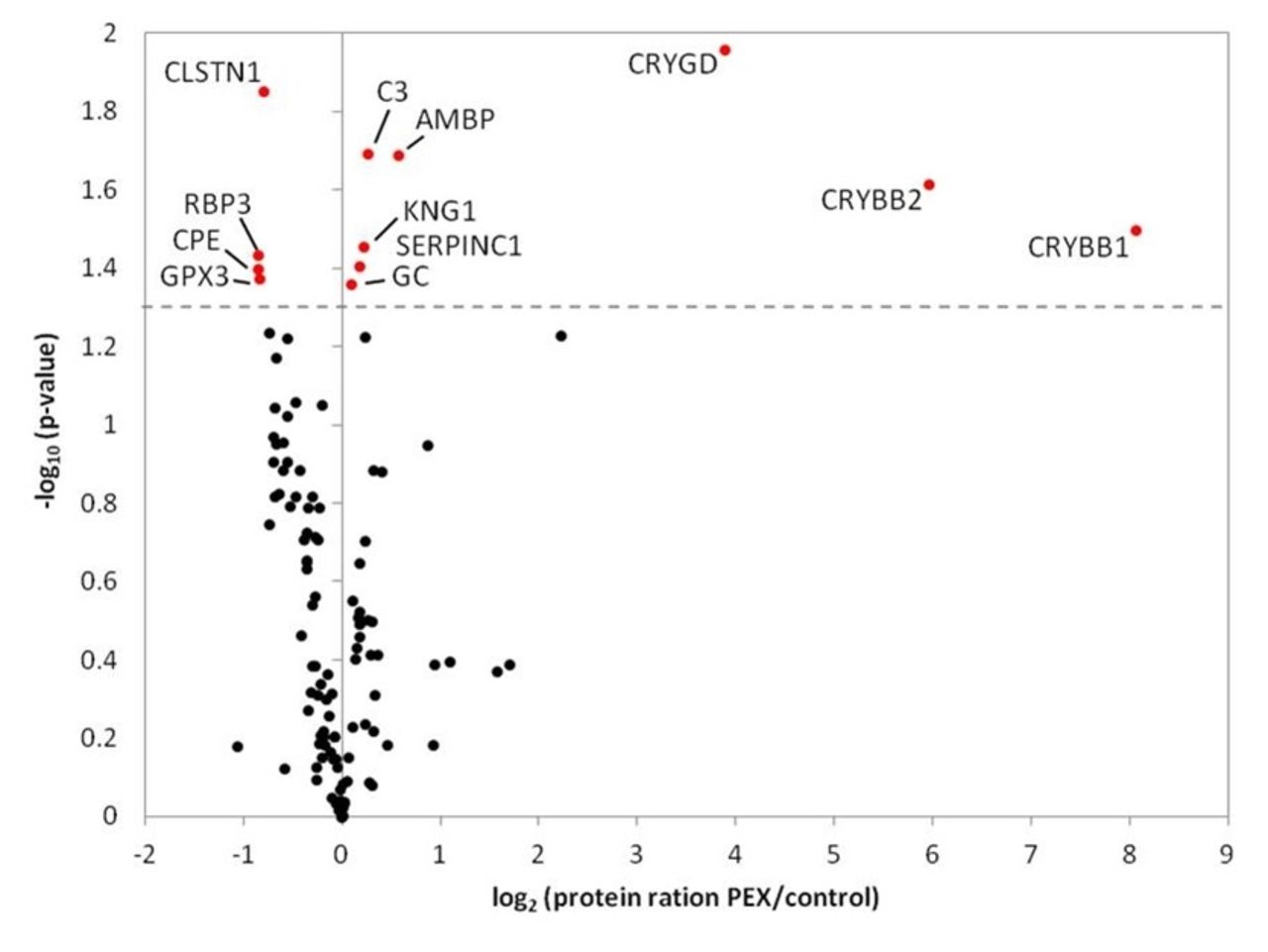
Protein quantification of PEX and control samples based on stable isotope dimethyl labelling.Volcano plot includes 113 proteins which were quantified in at least 4 out of 10 replicates. Protein rations (log2) were plotted against negative logarithmised p values (−log10). Significantly regulated proteins (p<0.05) are colouredin red and included CRYGD, CRYBB2, CRYBB1 and protein AMBP in addition to the proteins identified in figure 2. CRYBB1, beta-crystallin B1; CRYBB2, beta-crystallin B2; CRYGD, gamma-crystallin D; PEX, pseudoexfoliation syndrome.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Strong upregulation of crystallins in PEX samples. The intensities (log10) of all quantified proteins resulting from control (ctrl) samples are equivalent to all quantified proteins of PEX samples (left panel). CRYBB1, CRYBB2 and CRYGD were quantified with a significant and strong upregulation in PEX samples as illustrated based on the difference in detected protein intensities (right panel). Box plots representing median values ± ranges. Open circles are significant outliers. CRYBB1, beta-crystallin B1; CRYBB2, beta-crystallin B2; CRYGD, gamma-crystallin D; PEX, pseudoexfoliation syndrome.
All raw data from the MaxQuant searches are provided as pdf files in online supplemental material Suppl 1.1-Suppl 1.11.
Discussion
On the whole, with our global proteome analysis of AH of individuals with PEX, we managed to identify more than 200 proteins including candidates with significant regulation. In the present study, four proteins were increased in 10 out of 10 patients with PEX; C3, KNG1, antithrombin III and GC. The proteins RBP3, GPX3, CPE and CLSTN1 were decreased in 8 out of 10 of the individuals with PEX. These proteins are involved in various processes such as oxidative stress, inflammation and coagulation. The coagulation and immune systems are two host defence systems with complementary roles. Tissue damage, invading pathogens leads to activation of coagulation, and coagulation affects inflammatory response.8
C3 is a central part of the immune system. In the eye, C3 has been identified in PEX material previously, and lower concentrations of C3 in serum were found in patients with PEX glaucoma compared with patients with primary open angle glaucoma (POAG).9 10 The high molecule weight kininogen, HNK3, functions as a cofactor in the coagulation cascade and in the plasma bradykinin forming cascade, initiated by tissue damage, resulting in inflammatory response. Antithromibin III has been identified in human AH, trabecular meshwork, Schlemm’s canal and lens epithelium previously. Antithrombin III inhibits thrombin and formation of blood clots and has anti-inflammatory properties.11 GC is a plasma protein with various functions, such as regulation of vitamin D, transportation, bone remodelling and modulation of immune and inflammatory responses.12 RBP3 is synthesised in the ciliary epithelium, secreted into the AH and acts as a transportation protein in the retinal pigment epithelium.13 The concentration of RBP3 in AH is decreased in AH during autoimmune uveitis (AU) and treatment of AU restores the concentration of RBP3 to normal concentration.14 The decreased concentration of RBP3 and the increased concentration of C3 in AH in eyes with PEX indicate an inflammatory process. The multifunctional proteins HNK3, antithrombin III and GC are also connected with the immune system. The mechanism for their increase in the context PEX is unclear and remains to be elucidated.
GPX3 reduces hydrogen peroxide (H2O2) and other reactive oxygen species (ROS) to water by oxidising glutathione (GSH) to glutathione disulphide (GSSG). Previous studies indicate that oxidative stress is part of the pathogenesis in PEX. Gartaganis et al found a decreased concentration of GSH and an increased concentration of GSSG in eyes with PEX, and Ferreira et al described an increased activity of GPX3 in eyes with PEX glaucoma compared with eyes with POAG and controls.4 15 In the present study, the decreased GPX3 indicates a depletion of the enzyme by having an excess of ROS in the AH, thus confirming previous studies.
CPE is involved in the biosynthesis of peptide hormones and has a neuroprotective role in central nervous system by protecting neural cells from apoptosis when exposed to as oxidative stress or behavioural stress.16 In the eye, CPE has been identified in ciliary epithelium, photoreceptors and in ganglion cells in the retina. The expression of CPE in the retina decreases with age.17 One can hypothesise that a decreased level of CPE in eyes with PEX might indicate a deficient response to oxidative stress in these eyes. Calsyntenin is a calcium-binding postsynaptic membrane protein abundant in the brain.18 Gao et al investigated the vitreous in eyes with proliferative diabetes retinopathy and found that CLSTN1 and RBP3 were decreased compared with eyes without diabetes.19 CLSTN1 has been identified in AH previously, but the function in PEX is unclear.5
In a previous study, we identified increased concentrations of crystallins in a pooled material of AH from individuals with PEX. The PEX group had considerably increased concentrations of CRYBB1, CRYBB2 and CRYGS compared with controls.5 Therefore, it was of interest to investigate the content of crystallins in the present study. CRYBB1, CRYBB2 and CRYGD had up to eightfold increased concentrations in PEX in four of the PEX–control pairs. CRYBB2 was significantly increased in 5 out of 10 samples with PEX. In the remaining PEX–control pairs, the crystallins were below detection level which might indicate a subdivision within individuals with PEX.
The beta- and gamma-crystallins are structural proteins of the lens and have calcium-binding properties.20 In the lens, the crystallins undergo constant modifications with increasing age, affecting the structure and functions of the crystallins. These modifications can result in protein aggregation, light scattering and diminished transparency of the aged lens.21 Outside the lens, the functions of the crystallins are not clear, but several reports indicate changes in their expression during oxidative stress and inflammation. For example, the concentration of CRYBB2 was increased in the vitreous in eyes with uveitis.22 Sakaguchi et al reported increased content of crystallins in the retina after intense light exposure, and Yoshimura et al reported increased expression of crystallins following ischaemia-reperfusion injury.23 24 Fischer et al found promoted axon outgrowth, induced expression of ciliary neutrophic factor in astrocytes and Mϋller cells, and inflammatory reactions with infiltration of macrophages in the retina after injection of beta-/gamma-crystallin in the vitreous.25 One can speculate over the increased concentration of CRYBB1, CRYBB2B2 and CRYGD in the present study. An elevation of these crystallins in AH might represent a part of a stress-response reaction in the anterior segment in eyes with PEX.
Conclusions
Our results show that hundreds of proteins can be identified in small samples of body fluids such as AH in individual patients by using high-resolution mass spectrometry. Increased levels of C3, KNG-1, antithrombin III and GC and decreased levels of RBP3, CPE, GPX3 and CLSTN1 were identified in AH in eyes with PEX. The decrease in GPX3 and increase in C3 are in agreement with previous studies. The other proteins have multiple functions in the immune system and the coagulation cascade. Their functions in the eye are not clear in the context of PEX, but it is reasonable to assume that inflammation to some degree is involved in the pathogenesis of PEX.
Supplemental material
Supplemental material
Supplemental material
Supplemental material
Acknowledgments
The Swedish Research Council (2015-4870 (JB)), the foundation of Hanna Roos af Hjelmsäter, Uppsala and Stockholm County Councils (ALF project) are hereby acknowledged.
References
Footnotes
Contributors ABT collected the patient samples, ABT, AK and JB analysed the data, ABT AA and JB planned and conducted the study. All authors helped writing the paper.
Funding This study was funded by Vetenskapsrådet (grant no: 621-2011-4423, 2015-4870).
Competing interests None declared.
Patient consent for publication Obtained.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement All data are provided in the supplemental material.
Linked Articles
- At a glance