Article Text

Abstract
Aim To elucidate the role of microRNA-20a-5p (miR-20a-5p) in the pathogenesis of Vogt-Koyanagi-Harada (VKH) disease.
Methods Quantitative real-time PCR was used to quantify miR-20a-5p expression in CD4+ T cells from patients with active VKH and normal controls. The promoter methylation status of miR-20a-5p was detected by bisulfite sequencing PCR. Targets were evaluated by a luciferase reporter assay. The functional effects of miR-20a-5p on CD4+ T cells from patients with active VKH were assessed by upregulation or downregulation of its expression using liposomes.
Results The miR-20a-5p level was significantly decreased in CD4+ T cells from patients with active VKH as compared with normal controls. The two genes, oncostatin M (OSM) and C-C motif chemokine ligand 1 (CCL1), were identified as targets of miR-20a-5p. The upregulation of miR-20a-5p significantly suppressed interleukin 17 (IL-17) production in CD4+ T cells from patients with active VKH, whereas downregulation of miR-20a-5p exhibited an inverse effect. In addition, overexpression of OSM and CCL1 could rescue the effect of the upregulation of miR-20a-5p. Moreover, the level of miR-20a-5p was reduced in response to hypermethylation of the promoter. Further study showed that miR-20a-5p suppressed the activity of the phosphoinositide 3-kinase-AKT pathway.
Conclusions Our findings indicate that downregulation of miR-20a-5p is caused by promoter hypermethylation. MiR-20a-5p could also suppress the production of IL-17 by targeting OSM and CCL1 production in CD4+ T cells in patients with active VKH.
- experimental laboratory
- immunology
- inflammation
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Introduction
In China, Vogt-Koyanagi-Harada (VKH) disease is a common uveitis entity which is a great threat to vision. It is a multisystemic autoimmune disease directed against melanocyte-associated antigens and is characterised by a bilateral granulomatous panuveitis and extraocular symptoms including headache, scalp allergy, tinnitus, dysacusis, vitiligo, poliosis and alopecia.1 2 However, the exact pathogenesis of VKH disease remains elusive. It has been shown that HLA-DR4 and HLA-DRw53 are associated with VKH disease.3 4 Genetic variants located at 1p31.2 and 10q21.3 have also been shown to confer susceptibility to VKH disease.5 Growing evidence has demonstrated that CD4+ T lymphocyte autoreactivity, especially hyperactivity of Th17 and Th1 cells, plays an important role in the pathogenesis of VKH disease.6 7
MicroRNAs (miRNAs) are non-coding RNAs that range from 20nt to 24nt in terms of length and are capable of regulating post-transcriptional expression of genes, causing their degradation or suppressing mRNA translation. A large number of studies indicate that miRNAs are associated with various human diseases.8 9 Aberrant miRNA expression profiles may, for example, contribute to inflammatory bowel disease pathogenesis,10 and miR-126 regulates DNA methylation in CD4+ T cells by targeting DNA methyltransferase 1,11 12 which is involved in the pathogenesis of systemic lupus erythematosus and rheumatoid arthritis. A previous study from our group demonstrated that downregulation of miR-155 was involved in Behçet’s disease pathogenesis,13 and miR-146a was also associated with this disease.14 Recently, our team also found that miR-301a, miR-146a and miR-23a could confer susceptibility to VKH disease.15 These results provide a novel approach for investigating the role of miRNAs in the pathogenesis of uveitis.
Next-generation sequencing and microarray analysis showed that the expression of microRNA-20a-5p (miR-20a-5p) was decreased in multiple sclerosis (MS).16 There was a report showed that miR-20a-5p could be used as biomarker in the treatment response of naive patients with relapsing remitting MS with interferon-β (IFN-β).17 These findings also suggested that miR-20a-5p might be an important player in the pathophysiology of MS.18 Since the role of miR-20a-5p in uveitis is not completely clarified, we determined to investigate the expression pattern and role of this molecule in VKH disease. Our results demonstrated that miR-20a-5p was decreased in CD4+ T cells from patients with VKH with active intraocular inflammation and was downregulated due to the hypermethylation of its promoter. Moreover, it could suppress interleukin 17 (IL-17) production by inhibiting oncostatin M (OSM), C-C motif chemokine ligand 1 (CCL1) expression and the activity of the phosphoinositide 3-kinase (PI3K)-AKT pathway.
Materials and methods
Clinical specimens
Thirty-eight patients with VKH with active intraocular inflammation who had not taken any immunosuppressive drugs for at least 7 days (online supplementary table) and 25 normal controls were enrolled. The diagnosis of VKH disease was made in accordance with the diagnostic criteria for VKH disease by an international committee on nomenclature. All patients were diagnosed in our uveitis centre between March 2016 and April 2017, and they all showed ocular inflammation. All the subjects examined provided written informed consent before blood collection. The study was approved by the ethical committee of Chongqing Medical University. The tenets of the Declaration of Helsinki were followed during all procedures of the present study.
CD4+ T cell isolation
Ficoll-Hypaque density-gradient centrifugation was used to isolate peripheral blood mononuclear cells (PBMCs) from fresh heparinised blood of all the subjects in the study. CD4 mAb-conjugated magnetic microbeads (Miltenyi Biotec) were adopted to purify CD4+ T cells from PBMCs following its protocol and used directly or stored at −80°C for further study.
RNA preparation and real-time PCR
TRIzol (Invitrogen) was used to extract RNA, including miRNA. The levels of miRNAs were measured by quantitative real-time PCR using miDETECT A Track Kit (RiboBio) and performed on the ABI 7500 System (Applied Biosystems). The primers for miR-20a-5p and U6 small nuclear RNA were obtained from RiboBio Company, using sequences that cannot be disclosed because they are covered by a company-owned patent. Analyses of miR-20a-5p expression were normalised to the expression of internal control U6 using the 2−ΔΔCT method.
Transfection of miRNA mimics and inhibitor in CD4+ T cells
CD4+ T cells from patients with active VKH were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin and then seeded in 24-well plates of 1×106 cells/well; the cells were then transfected with miR-20a-5p mimics or inhibitor (RiboBio) using riboFECT CP Transfection Kit (RiboBio). Negative control mimics or inhibitor served as matched controls. To detect the effect of miR-20a-5p mimics and inhibitor on cytokine production by CD4+ T cells, the cells were stimulated with anti-CD3 (Miltenyi Biotec, 1 µg/mL) and anti-CD28 (Miltenyi Biotec, 1 µg/mL) and cultured for 72 hours at 37°C in 5% CO2. The concentration of IL-17 and IFN-γ in the culture supernatants was detected by ELISA.
OSM and CCL1 expression plasmid cotransfection with miR-20a-5p in CD4+ T cells
In our so-called rescue experiments, where we prepared miR-20a-5p/OSM and miR-20a-5p/CCL1 coexpressing cells, CD4+ T cells from patients with active VKH were seeded in 24-well plates (1×106 cells/well) with 500 µL medium. A mixture of miR-20a-5p mimics was then prepared using the riboFECT CP Transfection Kit following the manufacturer’s protocol. A mixture of OSM or CCL1 was prepared using Lipofectamine 3000 according to the manufacturer’s instructions. The mixture of miR-20a-5p mimics was added to the 24-well plates, whereafter the OSM–lipid or CCL1–lipid complex was added to the cells. Coexpressing cells were incubated for 3 days at 37°C in 5% CO2 and then analysed.
Flow cytometry
To stain intracellular cytokine, the CD4+ T cells from patients with active VKH were stimulated for 5 hours with PMA (MultiSciences, 50 ng/mL), ionomycin (MultiSciences, 1 µg/mL) and Brefeldin A (MultiSciences, 8 µg/mL) in the condition of 37°C and 5% CO2. Cells fixation and permeabilisation were performed using Cytofix/Cytoperm reagents (eBioscience) followed by staining antihuman IFN-γ (BD Biosciences) and antihuman IL-17A (BD Biosciences). Flow cytometry was carried out with a FACS Aria cytometer (BD Biosciences). The results were analysed by FlowJo software (Treestar).
Enzyme-linked immunosorbent assay
A human Duoset ELISA development kit (R&D Systems) was adopted to measure the concentration of IFN-γ and IL-17 from stimulated CD4+ T cells supernatants of patients with active VKH. ELISA Kits (Cloud-Clone) for OSM and CCL1 were used to measure OSM and CCL1 in cell culture supernatants.
DNA methylation analyses
Genomic DNA purification kit (Promega) was employed to obtain genomic DNA from CD4+ T cells of patients with active VKH and controls. The EpiTect Bisulfite Kit (Qiagen) was applied for DNA bisulfite modification. (F: 5′-GGTTGGGGGATATAAAGGAG-3′; R: 5′-ACCCGAAACTACAAAATACCC-3′) served as primers. Bisulfite sequencing PCR (BSP) was used to detect the promoter status. The PCR reaction was performed for 5 min at 95°C, for 30 s at 95°C with 40 cycles, for 30 s at 60°C, followed by 30 s at 72°C. The products were identified by electrophoresis and then cloned into pMD19-T-vector (TaKaRa) and sequenced (IGE Biotechnology).
Drug treatment
CD4+ T cells from patients with active VKH were treated with or without a demethylation agent, 5-Aza-2′-deoxycytidine (Sigma-Aldrich) at a concentration of 10 µM as described previously.19 Another batch of CD4+ T cells from patients with active VKH was treated with the PI3K-AKT signalling pathway inhibitor LY294002 (Sigma-Aldrich) at a concentration of 10 µM according to previous research20 and then stimulated with anti-CD28 and anti-CD3 for 72 hours. Supernatants and cells were then collected for further assays.
Dual-Luciferase Reporter assays
For luciferase activities assessment, the 3′UTR segments of OSM, CCL1 and SEMA7A containing a putative miR-20a-5p binding site were augmented by PCR and then inserted into the PGL-Basic-vector. Mutant constructs specific for putative miR-20a-5p binding sites in OSM, CCL1 and SEMA7A 3′UTR were also generated using QuickChange Site-Directed Mutagenesis Kit (Angilent). The pGL3-miR-20a-5p-Luc reporter, pGL-TK and miR-20a-5p-expressing vector were cotransfected using Lipo3000 (Invitrogen). Dual-Luciferase Reporter Assay Kit (Promega) was applied to assess the activities of luciferase after transfection for 48 hours.
Immunoblot analyses
Proteins were extracted with lysis buffer and then quantified by the bicinchoninic acid (BCA) method (KeyGen Biotech). Lysates were diluted in sodium dodecyl sulfate (SDS) sample buffer prior to SDS polyacrylamide gel electrophoresis and then transferred to polyvinylidene difluoride (PVDF) membranes. PVDF membranes were immunoblotted overnight at 4°C with primary antibodies (Abcam), followed by the appropriate secondary antibodies (Abcam). The bands were exposed using Pierce ECL Western Blotting Substrate (Thermo Fisher Scientific). Gel densitometry (Bio-Rad Laboratories) was used to quantify immunoblot signals on exposed film.
Statistical analysis
Analyses were carried out by SPSS V.13.0 statistical software, and graphic output was created using GraphPad Prism V.5.0 software. Quantitative values of all experiments are presented as the mean±SD. Statistical significance among/between groups were determined by one-way analysis of variance or independent-samples Student’s t-test. Pearson’s correlation analysis was used to evaluate the correlation between miR-20a-5p and OSM, and miR-20a-5p and CCL1. p<0.05 was considered to be of statistically significance.
Results
Decreased miR-20a-5p contributes to increased production of IL-17 and frequencies of IL-17+ CD4+ T cells in patients with active VKH
For investigating the expression pattern of miR-20a-5p in VKH disease, we measured the level of miR-20a-5p in CD4+ T cells from 14 active patients and 14 controls. PCR results indicated that miR-20a-5p was significantly decreased in CD4+ T cells from patients with active VKH in comparison to normal controls (figure 1A). Former researches reported that Th17 and Th1 cells play a critical role in the inflammatory activity of VKH disease.7 21 To explore the possible effects of miR-20a-5p on Th1 or Th17 function, we transfected miR-20a-5p-expressing mimics or miR-20a-5p-repressing inhibitor into CD4+ T cells from patients with active VKH, respectively (online supplementary figure 1A). We analysed the frequencies of IL-17+ and IFN-γ+ CD4+ T cells in six patients with active VKH by flow cytometry. The results suggested that inhibition of miR-20a-5p upregulated intracellular IL-17 level in CD4+ T cells, while upregulation of miR-20a-5p yielded the opposite effect (figure 1B,C). We also examined the production of IFN-γ and IL-17 in the supernatants of CD4+ T cell and found that ectopic expression of miR-20a-5p reduced IL-17 content, as shown by ELISA, while silencing of miR-20a-5p showed the opposite effect (figure 1D,E). The frequency of IFN-γ+ CD4+ T cells and IFN-γ release was not affected when we altered the expression of miR-20a-5p (online supplementary figure 1B–E).

Downregulation of miR-20a-5p enhances frequencies of IL-17+ CD4+ T cells and IL-17 production in patients with active VKH. (A) miR-20a-5p expression was examined by quantitative real-time PCR in CD4+ T cells from 14 patients with active VKH and 14 normal controls. (B, C) Analysis of frequencies of IL-17+ CD4+ T cells in patients with active VKH by flow cytometry after treating CD4+ T cells with miR-20a-5p mimics or miR-20a-5p inhibitor. (D, E) Detection of IL-17 production in the supernatants by ELISA after treating CD4+ T cells from patients with active VKH with miR-20a-5p mimics or miR-20a-5p inhibitor. Independent-samples Student’s t-test was used for statistical analyses. IL-17, interleukin 17; miR-20a-5p, microRNA-20a-5p; NC, negative control; VKH, Vogt-Koyanagi-Harada.
Decreased miR-20a-5p is associated with promoter hypermethylation in patients with active VKH
To explore whether transcriptional silencing of miR-20a-5p is associated with aberrant promoter methylation, we used the UCSC Genome Browser to identify the miR-20a-5p promoter. It was located in the 5 kb region directly upstream of the miR-20a-5p stem loop. We then used MethPrimer to analyse the promoter methylation status and identified the CpG islands (see online supplementary figure 2A). The region from −1793 bp to −2041 bp of the stem loop of miR-20a-5p was amplified successfully, and BSP results showed that there was a higher methylation frequency of this site in five patients with active VKH in contrast to normal controls (figure 2A–C). In subsequent experiments, we exposed the CD4+ T cells from active patients to the methylation inhibitor 5-Aza-2′-deoxycytidine and results indeed showed a decreased methylation frequency (figure 2D, online supplementary figure 2B), which was associated with increased levels of miR-20a-5p in treated samples (figure 2E).
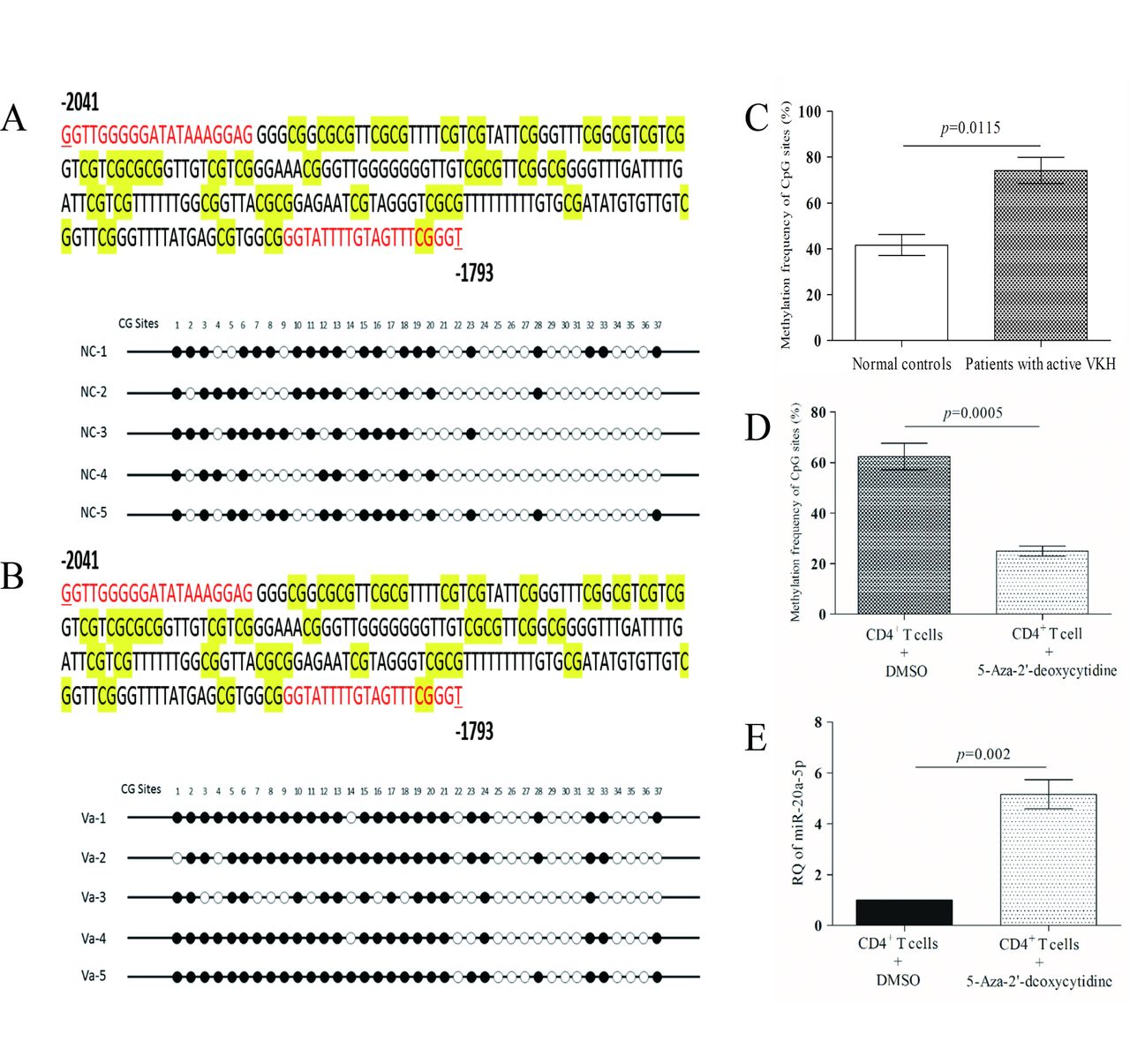
Promoter hypermethylation contributes to decreased expression of miR-20a-5p in CD4+ T cells from patients with active VKH. (A) DNA sequencing results of methylation sites in CD4+ T cells from normal controls. (B) DNA sequencing results of methylation sites in CD4+ T cells from patients with active VKH. (C) Methylation frequency of CpG sites of patients with active VKH as compared with that of normal controls. (D) Comparison of of the methylation frequency of CpG sites of CD4+ T cells between those treated with 5-Aza-2′-deoxycytidine and DMSO carrier in patients with active VKH. (E) Detection of miR-20a-5p expression after 5-Aza-2′-deoxycytidine treatment in CD4+ T cells from patients with active VKH. The relative expression level without 5-Aza-2′-deoxycytidine treatment was normalised to 1. Independent-samples Student’s t-test was used for statistical analyses. DMSO, dimethyl sulfoxide; miR-20a-5p, microRNA-20a-5p; VKH, Vogt-Koyanagi-Harada.
OSM and CCL1 are direct targets of miR-20a-5p
To identify effectors of miR-20a-5p, we used the bioinformatic algorithm named OmicsBean, which contains eight miRNA prediction databases (http://www.omicsbean.com:88/). Several target mRNAs were predicted to be regulated by miR-20a-5p. We then focused on mRNAs involved in immune and inflammation responses using biological process enrichment procedures (see online supplementary figure 3A,B). Among those mRNAs, OSM, CCL1 and SEMA7A were predicted, as shown in online supplementary figure 3C. We cloned the 3′UTRs of OSM, CCL1 and SEMA7A into a Dual-Luciferase vector. Luciferase activities assessments revealed that miR-20a-5p repressed the luciferase activity of wild-type OSM and CCL1 3′UTR in 293 T cells but it had no effect on the activity of mutant-type OSM and CCL1 3′UTR (figure 3A,B). Moreover, exogenous expression of miR-20a-5p significantly reduced the amount of OSM and CCL1 protein in CD4+ T cells (figure 3C) and supernatants of cultured medium from active patients as shown by immunoblotting and ELISA (figure 3D,E). Since the effect on CCL1 protein expression was weak, the experiment was repeated on a separate occasion and again showed a significant inhibitory effect of miR-20a-5p (data not shown). However, miR-20a-5p was not able to inhibit the luciferase activity of 3′UTR of SEMA7A (see online supplementary figure 3D).
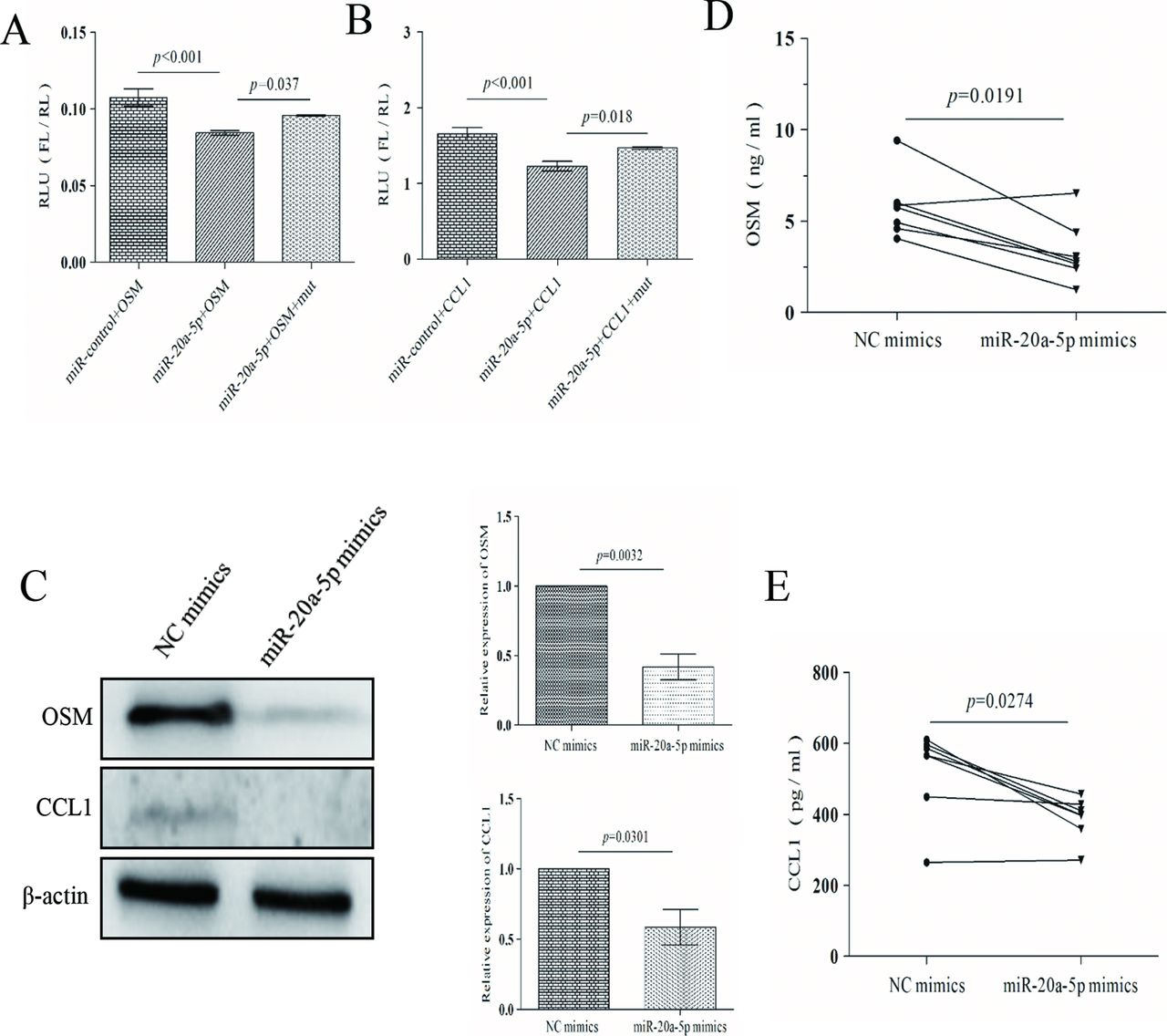
OSM and CCL1 are direct targets of miR-20a-5p. (A) Luciferase activities of wild-type and mutant-type 3′UTR of OSM in 293 T cells, after transfection of miR-20a-5p mimics. (B) Luciferase activities of wild-type and mutant-type 3′UTR of CCL1 in 293 T cells, after transfection of miR-20a-5p mimics. (C) OSM and CCL1 expression in CD4+ T cells from patients with active VKH treated with miR-20a-5p mimics by western blot (n=3). (D) Detection of OSM production in the supernatants by ELISA after treating CD4+ T cells from patients with active VKH with miR-20a-5p mimics. (E) Detection of CCL1 production in the supernatants by ELISA after treating CD4+ T cells from patients with active VKH with miR-20a-5p mimics. Independent-samples Student’s t-test for two independent samples and ANOVA for multigroup samples were used for statistical analyses. ANOVA, analysis of variance; CCL1, C-C motif chemokine ligand 1; FL, firefly luciferase; miR-20a-5p, microRNA-20a-5p; OSM, oncostatin M; RL, renilla luciferase; RLU, relative luciferase activity; VKH, Vogt-Koyanagi-Harada.
OSM and CCL1 are functional mediators of miR-20a-5p and show inverse correlation with miR-20a-5p in VKH disease
To investigate the effects of OSM and CCL1, we established OSM and CCL1 overexpression vectors not containing 3′UTR (see online supplementary figure 4). As shown in figure 4A,B, overexpression of OSM increased the frequencies of IL-17+ CD4+ T cells and supernatant IL-17 production of CD4+ T cells from patients with active VKH, and overexpression of CCL1 showed the same effects.

OSM and CCL1 are functional targets of miR-20a-5p in VKH disease. (A) Detection of IL-17 production in the supernatants by ELISA after transfecting CD4+ T cells from patients with active VKH with OSM and CCL1 overexpression vectors. (B) Analysis of frequencies of IL-17+ CD4+ T cells in patients with active VKH by flow cytometry after transfecting CD4+ T cells with OSM and CCL1 overexpression vectors. (C) Analysis of frequencies of IL-17+ CD4+ T cells in patients with active VKH by flow cytometry after treating CD4+ T cells with miR-20a-5p mimics and miR-20a-5p/OSM and miR-20a-5p mimics and miR-20a-5p/CCL1. (D, E) Detection of IL-17 production in the supernatants by ELISA after treating CD4+ T cells from patients with active VKH with miR-20a-5p mimics and miR-20a-5p/OSM (D) and miR-20a-5p mimics and miR-20a-5p/CCL1 (E). Independent-samples Student’s t-test for two independent samples and ANOVA for multigroup samples were used for statistical analyses. ANOVA, analysis of variance; CCL1, C-C motif chemokine ligand 1; IL-17, interleukin 17; miR-20a-5p, microRNA-20a-5p; OSM, oncostatin M; VKH, Vogt-Koyanagi-Harada.
Based on the results of Dual-Luciferase assays, we hypothesised that OSM and CCL1 mediated the effects of miR-20a-5p. To test it, the miR-20a-5p mimics and OSM overexpression vectors not containing 3′UTR (miR-20a-5p/OSM) and miR-20a-5p mimics and CCL1 overexpression vectors not containing 3′UTR (miR-20a-5p/CCL1) were transfected into CD4+ T cells from patients with active VKH, respectively (see online supplementary figure 4B). Results demonstrated that increased miR-20a-5p expression reduced frequencies of IL-17+ CD4+ T cells (figure 4C) and supernatant (figure 4D,E) IL-17 production of CD4+ T cells from patients with active VKH, while miR-20a-5p/OSM and miR-20a-5p/CCL1 could reverse miR-20a-5p upregulation-imposed effects, at least partly. (figure 4C–E). Moreover, we measured miR-20a-5p, OSM and CCL1 expression in five patients with active VKH and five normal controls and found that the expression level of miR-20a-5p was lower in patients with active VKH than that in normal controls (figure 5A), while OSM and CCL1 showed the opposite expression profiles (figure 5B,C). Pearson correlation analysis showed that there was an inverse correlation between miR-20a-5p and OSM protein level (r=−0.719, p=0.019; figure 5D) and miR-20a-5p and CCL1 protein level (r=−0.651, p=0.042; figure 5D).

miR-20a-5p is inversely correlated with OSM and CCL1 in patients with active VKH. (A, B) Detection of miR-20a-5p, OSM and CCL1 expression levels. (A) miR-20a-5p was examined by quantitative real-time PCR, and (B) OSM and CCL1 protein levels were detected by western blot in CD4+ T cells from five patients with active VKH and five normal controls. (C) The relative protein levels of OSM and CCL1 were analysed by Fusion software, and their expression levels were normalised to expression level of N1. (D) Inverse correlation of miR-20a-5p expression with OSM protein level and with CCL1 protein level in CD4+ T cell from patients with active VKH (n=5) and normal controls (n=5). CCL1, C-C motif chemokine ligand 1; miR-20a-5p, microRNA-20a-5p; OSM, oncostatin M; RQ, relative quantification; VKH, Vogt-Koyanagi-Harada.
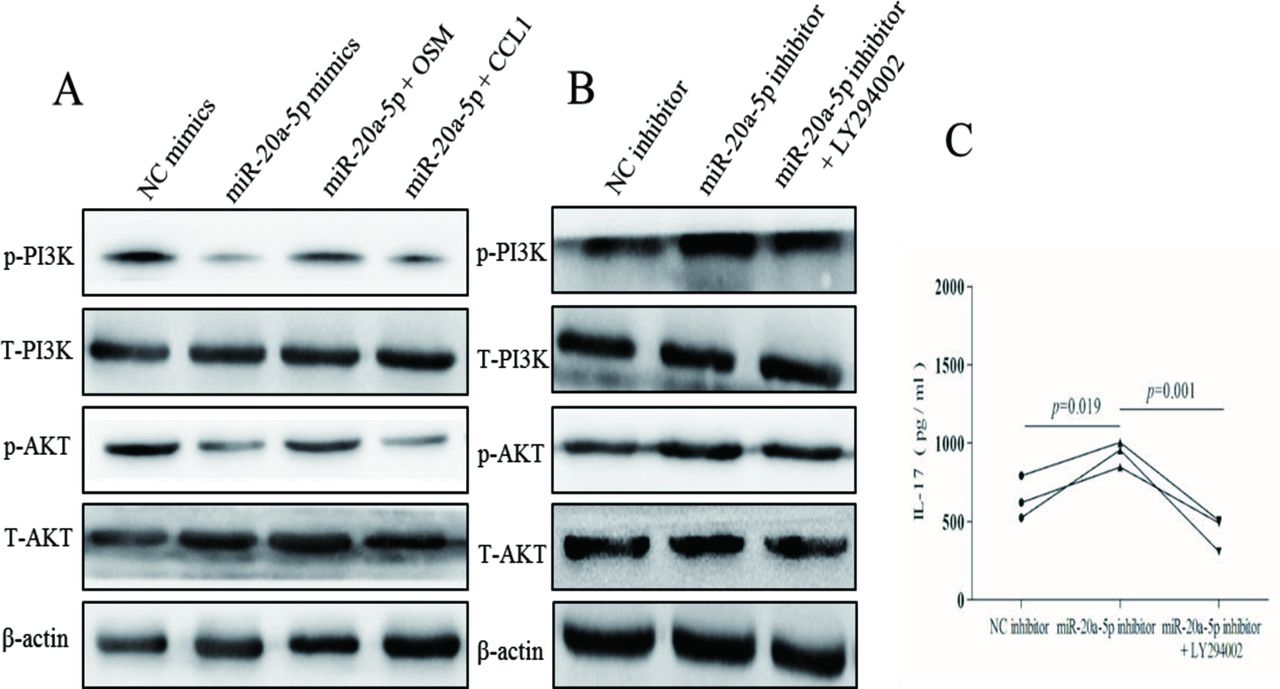
miR-20a-5p is involved in PI3K-AKT signalling pathways in VKH disease
Previous research has shown that PI3K-AKT pathway is associated with several immune and inflammatory diseases22 23 and that some miRNAs could regulate this pathway.24 25 Since this has not yet been studied in uveitis, we wondered whether miR-20a-5p participates in the pathogenesis of VKH disease via the PI3K-AKT pathway. Western blot analysis showed reduced phosphorylation levels of AKT and PI3K after overexpressing miR-20a-5p (figure 6A) and vice versa (figure 6B). Interestingly, miR-20a-5p/OSM obviously rescued the inhibitory effect of miR-20a-5p on p-PI3K and p-AKT; however, miR-20a-5p/CCL1 could not exhibit rescue effect (figure 6A). The total protein amount of AKT and PI3K was not changed under any circumstance (figure 6A,B). We treated CD4+ T cells from patients with active VKH with miR-20a-5p inhibitor transfection and then with PI3K-AKT inhibitor LY294002 for 3 days. The results showed that PI3K-AKT inhibitor LY294002 could decrease p-PI3K and p-AKT activities enhanced by miR-20a-5p inhibitor (figure 6B). We also found that LY294002 could abrogate the upregulation of IL-17 protein in miR-20a-5p-inhibitor-containing cells (figure 6C).

![[SP2.jpg]](https://bjo.bmj.com/content/bjophthalmol/102/2/282/DC2/embed/inline-supplementary-material-2.jpg?download=true){kind=link}
{kind=link}
![[SP3.jpg]](https://bjo.bmj.com/content/bjophthalmol/102/2/282/DC3/embed/inline-supplementary-material-3.jpg?download=true){kind=link}
{kind=link}
![[SP4.jpg]](https://bjo.bmj.com/content/bjophthalmol/102/2/282/DC4/embed/inline-supplementary-material-4.jpg?download=true){kind=link}
{kind=link}
![[SP5.jpg]](https://bjo.bmj.com/content/bjophthalmol/102/2/282/DC5/embed/inline-supplementary-material-5.jpg?download=true){kind=link}
{kind=link}
{kind=link}
{kind=link}
miR-20a-5p is a PI3K-AKT signalling pathway regulator in active VKH. (A) Analyses of p-PI3K, T-PI3K, p-AKT and T-AKT proteins level in CD4+ T cells from active VKH by western blot after miR-20a-5p mimics, miR-20a-5p/OSM and miR-20a-5p/CCL1 treatment. (B) Analyses of p-PI3K, T-PI3K, p-AKT and T-AKT proteins level in CD4+ T cells from active VKH by western blot after miR-20a-5p inhibitor and miR-20a-5p inhibitor treatment. (C) Detection of IL-17 production in the supernatants of CD4+ T cells from three patients with active VKH by ELISA after miR-20a-5p inhibitor and miR-20a-5p inhibitor treatment. ANOVA for multigroup samples was used for statistical analyses. ANOVA, analysis of variance; CCL1, C-C motif chemokine ligand 1; IL-17, interleukin 17; miR-20a-5p, microRNA-20a-5p; OSM, oncostatin M; VKH, Vogt-Koyanagi-Harada.
Discussion
This study demonstrated that miR-20a-5p is significantly decreased in CD4+ T cells from patients with active VKH as compared with normal controls. We also showed that miR-20a-5p suppressed the production of the proinflammatory cytokine IL-17 but did not alter production of IFN-γ, suggesting that a low miR-20a-5p is associated with an enhanced inflammatory response. A downregulation of miR-20a-5p has also been reported earlier in several autoimmune diseases17 and human cancer26 but has not yet been investigated during intraocular inflammation.
Promoter hypermethylation of miRNAs has been shown to be associated with their reduced expression.27 We confirmed these previous findings and showed that the promoter of miR-20a-5p from patients with active VKH showed a higher methylation frequency compared with normal controls. This was further strengthened by the finding of a decreased methylation frequency and increased mRNA levels of miR-20a-5p following treatment with the 5-Aza-2′-deoxycytidine.
To investigate the potential mechanisms explaining the roles of miR-20a-5p in pathogenesis of VKH disease, we analysed downstream targets of miR-20a-5p and identified OSM and CCL1 as functional effectors of miR-20a-5p. OSM belongs to the interleukin 6 family, which plays a role in the inflammatory response.28 Recently, it has been found to be involved in the expression of transcription factors associated with inflammatory and immune responses mediated by T cells.29 OSM has been considered as a proinflammatory cytokine,30 which is consistent with our findings, though it also been shown to exhibit anti-inflammatory functions.31 CCL1 is one of several chemokine genes clustered on the q-arm of chromosome 17.32 This chemokine is a member of the CC subfamily and is secreted by activated T cells and binds to the chemokine C-C motif receptor 8.33 It has been shown to be involved in immunoregulatory and inflammatory processes.34 35 Our results showed that OSM and CCL1 had proinflammatory activity in patients with VKH disease, which is probably mediated by increasing IL-17 production. Interestingly, previous studies show that IL-17 has synergistic effects with OSM on regulating skin inflammation,36 human rheumatoid arthritis37 and cartilage collagen breakdown.38 Importantly, we found that OSM and CCL1 could counteract the inhibitory effects of miR-20a-5p on the production of IL-17 in patients with active VKH, which supports the hypothesis that OSM and CCL1 are functional targets of miR-20a-5p, which in turn may affect the pathogenesis of VKH disease.
Having established the possible role of miR-20a-5p as an inflammation suppressor miRNA in VKH disease, we investigated possible pathways by which miR-20a-5p could be participated in this disease. The PI3K-AKT pathway plays an important role in several autoimmune diseases, and it has been shown that miRNAs may regulate this pathway.22 24 25 Our findings showed that miR-20a-5p mimics resulted in decreased phosphorylation levels of AKT and PI3K and vice versa. Importantly, the increased production of IL-17 induced by miR-20a-5p inhibitor was dramatically dampened after treatment with LY294002, a PI3K-AKT pathway inhibitor. These results suggested that the PI3K-AKT pathway contributed to IL-17 production and might be involved in controlling the effect of miR-20a-5p on the pathogenesis of VKH disease. Consistent with our findings, previous studies in mice reported that the PI3K-AKT pathway promotes Th17 cell differentiation and IL-17 production.39 Similarly, in experimental autoimmune encephalomyelitis,40 PI3K drives the disease pathogenesis by promoting Th17 differentiation. Conversely, inhibition of PI3K reduces IL-17 expression in a murine asthma model41 and psoriasis-like dermatitis.42 We found that overexpression of OSM was sufficient to rescue the decreased activities of the PI3K-AKT pathway by miR-20a-5p mimics, suggesting that OSM was involved in regulating the activity of the PI3K-AKT pathway. This might provide a mechanism to explain that OSM promotes IL-17 production in VKH disease. Consistently, it has been reported that OSM could protect against cardiac ischaemia/reperfusion injury in diabetic mice by activating the PI3K-AKT pathway.43 Of interest is the observation that OSM expression can be induced by the PI3K-AKT pathway in osteoblasts.20 44 This suggests a regulatory loop between OSM and the PI3K-AKT pathway. Unexpectedly, the rescue effect was not observed in the miR-20a-5p/CCL1 group.
To conclude, we propose a novel role for miR-20a-5p in the pathogenesis of VKH disease, wherein hypermethylation of the promoter contributed to downregulated expression of miR-20a-5p, which suppressed its targets OSM and CCL1. We found that miR-20a-5p reduced the production of IL-17 in CD4+ T cells from patients with active VKH. Moreover, miR-20a-5p could inhibit the activity of the PI3K-AKT pathway. Identification of inflammation-specific miRNAs and their targets is critical for understanding their role in the pathogenesis of VKH disease and may be used in the future development of drugs to treat this disease.
Acknowledgments
We thank all the volunteers who provided samples for the analyses presented in this study. We also thank all donors enrolled in the present study.
References
Footnotes
Contributors RC, PY: conceived the idea and designed the experiments. CZ, QC: sample collection. RC, SY, QW: performed all the experiments. GY: flow cytometry test. RC, YH: analysed the data. RC, XT: wrote the manuscript. PY, GS, AK: revised the manuscript.
Funding This work was supported by National Key R&D Program of China (grant no 2016YFC0904000), Natural Science Major International (Regional) Joint Research Project (grant no 81320108009), National Natural Science Foundation Project (grant no 31370893), Chongqing Key Laboratory of Ophthalmology (grant no CSTC, 2008CA5003), National Key Clinical Specialties Construction Program of China, Chongqing Science & Technology Platform and Base Construction Program (grant no cstc2014pt-sy10002) and National Natural Science Foundation Project (grant no 81400390).
Competing interests None declared.
Patient consent Obtained.
Ethics approval The study was approved by the ethical committee of Chongqing Medical University. The tenets of the Declaration of Helsinki were followed during all procedures of the present study.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- At a glance