Article Text

Abstract
Purpose This study examines the effect of the L-type calcium channel blocker verapamil on mechanical strain-induced extracellular matrix genes in optic nerve head lamina cribrosa (LC) cells.
Methods Changes in LC cell intracellular calcium [Ca2+]i following hypotonic cell membrane stretch were measured with the fluorescent probe fura-2/AM. Fluorescence intensity was measured, after labelling, by calcium (Ca2+) imaging confocal microscopy. Confluent human LC cell cultures were serum starved for 24 h prior to exposure to cyclical mechanical strain (1 Hz, 15%) for 24 h in the presence or absence of verapamil (10 mm). Transforming growth factor-β 1 (TGF-β1), collagen 6A3 (COL6A3) and chondroitin sulfate proteoglycan 2 (CSPG2) mRNA expression levels were assessed by quantitative RT-PCR.
Results Hypotonic cell membrane stretch of LC cells from normal donors significantly increased [Ca2+]i (p<0.05). Exposure to cyclical mechanical strain (15% strain) produced a statistically significant increase in the three matrix genes that were examined (TGF-β1, COL6A3 and CSPG2). This response in both cyclical and mechanical stretch was significantly reduced by pretreating LC cells with the L-type calcium channel blocker verapamil (p<0.05).
Conclusions This study provides evidence of a novel mechanotransduction pathway linking mechanical strain, cation channel function and the induction of LC cell matrix gene transcription. This highlights the potential involvement of calcium influx in the activation of matrix remodelling responses in the optic nerve head and supports the rationale that calcium channel blockers may attenuate disease progression in glaucoma.
- Glaucoma
Statistics from Altmetric.com
Introduction
Glaucoma is a chronic, degenerative optic neuropathy that can be distinguished from most other forms of acquired optic neuropathy by the characteristic appearance of the optic nerve.1 Within the cores of the cribriform plates in glaucoma, there is a marked decrease in the density of collagen fibrils around elastic fibres, altering the mechanical properties of the tissue during the glaucomatous process.2 Connective tissue remodelling in glaucoma alters the biomechanical response of the lamina cribrosa (LC) to intraocular pressure (IOP) elevation in an eye-specific manner.3 LC cells have a polygonal morphology and in vivo extend thin processes throughout the cribriform plates that make contact with adjacent astrocytes. They can be distinguished from astrocytes by absence of glial fibrillary acidic protein (GFAP) expression, in vivo or in vitro, and absence of vascular specific markers or other microglial markers.4
Mechanotransduction, by definition, refers to the mechanisms by which cells convert a mechanical stimulus into a chemical signal.5 Mechanical strain has been shown to induce a diverse array of biological effects including gene transcription, cell proliferation, differentiation and apoptosis.6 Abnormal mechanical strain on cells can alter the cellular function, change the composition of the extracellular matrix (ECM) and lead to organic pathology in atherosclerosis and interstitial lung disease, for example.7 The mechanisms by which altered intraocular forces induce a biological response are poorly understood, although a general mechanism for mechanotransduction has been proposed.8 Stretch-activated channels (SAC) and voltage-gated ion channels have both been reported to serve as mechanotransducers.9 Induction of SAC causes an intracellular accumulation of Ca2+ that can serve to trigger multiple signalling cascades and, ultimately, may contribute to remodelling of the ECM in response to strain.10 SAC channels have been isolated in many different cell types including LC cells.11
While Ca2+ is essential for a number of important cellular functions, an excessive elevation of intracellular calcium [Ca2+]i promotes pathological changes in a number of organs. The L-type Ca2+ channel blocker verapamil restricts the influx of Ca2+ across the cell membrane.12 L-type Ca2+ channels are expressed throughout the eye including the retinal pigment epithelium cells, ciliary body epithelium and the trabecular meshwork (TM).13 Persistent Ca2+ influx through these channels is often associated with prevalent pathology in the nervous, muscular and cardiovascular systems.14
He et al15 have implicated elevated [Ca2+]i and mitochondrial dysfunction in the pathological process that occurs in the TM of people with glaucoma. A previous study concluded that TM cells activate SAC causing an increase in [Ca2+]i.15 A study by our group similarly demonstrated elevated [Ca2+]i and abnormal calcium homeostasis mechanisms in LC cells from glaucomatous donors.16
Of relevance to our current study are recent works demonstrating that calcium channel blockers can significantly alter markers of fibrogenic signalling (including transforming growth factor- β1, TGF-β1), oxidative stress, IOP and optic nerve head circulation.17 ,18 TGF-β has been implicated in both the profibrotic response and the response to strain of LC cells19 and, interestingly, Ca2+ signalling has been implicated in the induction of TGF-β expression in a model of hepatic fibrosis.20
All three genes analysed in this study are upregulated in the glaucomatous optic nerve hypoplasia21 and have also been identified as strain responsive (at 15% level of strain) in LC cells previously.22 From our previous experimental models of strain, we were aware that 15% strain was the lowest level of strain needed to induce a significant ECM response using the Flexercell system.
In this paper, we seek to further investigate the relationship between strain and [Ca2+]i levels. Specifically, we analyse the response of [Ca2+]i to hypotonic strain by examining the response of the ECM genes COL6A3, chondroitin sulfate proteoglycan 2 (CSPG2) and TGF-β1 to the calcium channel blocker verapamil in the presence and absence of cyclical strain (15%) in LC cells.
Material and methods
LC cell isolation and culture
Eyes were obtained from the Lions Eye Institute for Transplant and Research, Tampa, Florida, USA. Isolation of LC cells was carried out on eyes received within 24 h of death, donors or a first degree relative having given consent for using the eyes for research purposes. LC cell explants from the same were subsequently donated by Alcon Laboratories, Fort Worth, Texas, USA. The acquisition of these cells was carried out in accordance with the Declaration of Helsinki.
The LC was carefully dissected and explanted. The cell lines used were characterised by immune fluorescence staining for a number of glial cell markers. Cells were deemed to be LC cells if they stained positively for elastin, fibronectin, laminin, α-smooth muscle actin and collagen I, III and IV, and if they stained negatively for GFAP. LC cells were also tested for GFAP by RT-PCR as per Lambert et al.23
LC cell lines from a total of five different donors with no history of glaucoma and ages ranging from 68 to 91 years, mean age 81.0±10.2 years, and from a total of four different donors with glaucoma and ages ranging from 68 to 83 years, mean age 77.8±6.4 years, were used in this study.
Primary cell culture lines of GFAP negative LC cells were cultured and maintained in Dulbecco's modified Eagle's medium supplemented with fetal bovine serum, penicillin (100 U/mL) and streptomycin (100 µg/mL) under atmospheric conditions of 5% CO2/21% O2 at 37°C, as previously described.21 ,22 For the Flexercell experiments, each cell line was seeded in triplicate onto three separate wells of a laminin coated Bioflex (Flexercell International Hillsborough, North Carolina, USA) stretch plate.
Calcium imaging
An increase in [Ca2+]i was measured using the Ca2+-sensitive fluorescent dye fura-2/AM (Bioscience, Molecular Probes, Ireland) as previously described.11 Cells were excited alternately at wavelengths of 340 and 380 nm. The resultant fluorescence at each excitation wavelength was measured at 510 nm collected using a charge-coupled devices camera system (Hammatsu, Japan). For Ca2+ calibration, values for [Ca2+]i were obtained from the following equation: where the dissociation constant (Kd) was assumed to be 225 nm based on the work of Grynkyiewicz et al24. Rmin is the ratio of fluorescence measured at 340 nm (F340) over 380 nm (F380) in a nominally Ca2+-free solution. Rmax is the ratio of fluorescence measured at 340 nm over 380 nm in the presence of saturating amounts of Ca2+ (10 mm CaCl2) and ionomycin (10 µm) in the solution. All calcium-imaging experiments were performed in dark at room temperature (20°C–22°C) to minimise dye leakage. LC cells were first settled in isotonic solution, then stimulated by a hypotonic solution in the presence of 2 mm extracellular Ca2+. Cells were either pretreated or non-treated (control) with verapamil (10 mM verapamil (Sigma)). Images were digitised and analysed using Openlab2 software (Improvision, UK).
where the dissociation constant (Kd) was assumed to be 225 nm based on the work of Grynkyiewicz et al24. Rmin is the ratio of fluorescence measured at 340 nm (F340) over 380 nm (F380) in a nominally Ca2+-free solution. Rmax is the ratio of fluorescence measured at 340 nm over 380 nm in the presence of saturating amounts of Ca2+ (10 mm CaCl2) and ionomycin (10 µm) in the solution. All calcium-imaging experiments were performed in dark at room temperature (20°C–22°C) to minimise dye leakage. LC cells were first settled in isotonic solution, then stimulated by a hypotonic solution in the presence of 2 mm extracellular Ca2+. Cells were either pretreated or non-treated (control) with verapamil (10 mM verapamil (Sigma)). Images were digitised and analysed using Openlab2 software (Improvision, UK).
Solutions
The isotonic solution contained (in mm) 120 NaCl, 6 KCl, 1 MgCl2, 2 CaCl2, 5.4 HEPES and 80 D-mannitol, pH 7.4 adjusted with NaOH (osmolality: 323±6 mOsm). The hypotonic solution (osmolarity: 232±8 mOsm) was prepared by omitting D-mannitol from the isotonic solution.
Agarose gel electrophoresis
The L-type calcium channel primer pair to amplify the α subunit of the channel was designed using Q primer-depot to ensure target gene specificity and spanning exon–exon junctions. The following primer pair was used for gene specific amplification of complimentary DNA (cDNA) from three LC cell strains:
Forward 5′-AAAGTTGGTGATGCCTCCAT-3′ Reverse 5′-AAAGTTGGTGATGCCTCCAT-3′
The products were visualised by 1% agarose gel electrophoresis stained with ethidium bromide. A 100 bp DNA ladder was used as a standard.
Application of mechanical strain
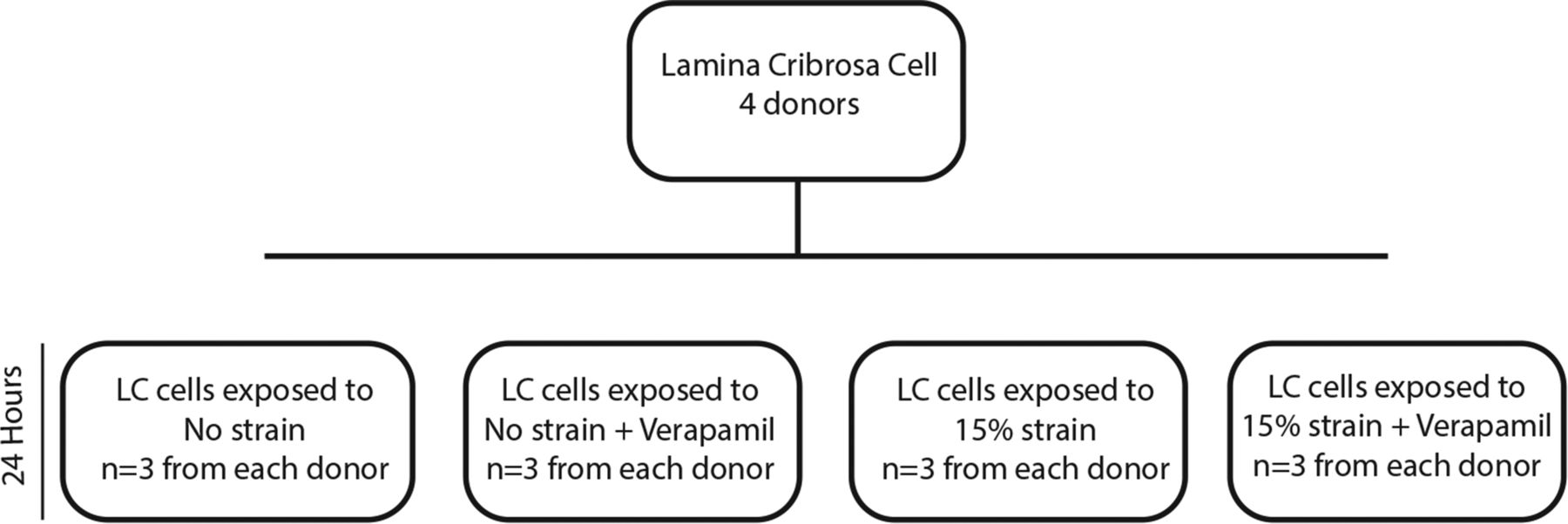
Triplicate cultures, from each donor, of confluent LC cells (four normal human donors) were grown on laminin coated Bioflex stretch plates and exposed to 15% cyclical strain at 1 Hz for 24 h in isotonic solutions using the Flexercell tension plus FX-4000 T system (Flexercell International, Hillsborough, North Carolina, USA). Triplicate controls for each donor were maintained in the same environment with no strain for 24 h. All cells were serum starved for 24 h prior to exposure to strain. One group of cells from each of the experimental conditions (0% and 15% strain) was treated with 10 mm verapamil (Sigma) 30 min prior to strain (figure 1).

Flexercell experimental setup. Lamina cribrosa (LC) cells were exposed to four experimental conditions for 24 h. Two groups were exposed to 15% strain and two groups were exposed to no strain. Verapamil (10 mM verapamil) was added 30 min prior to the start of the experiment to two groups, one at 15% strain and one at 0% strain.
RNA extraction
Total RNA (n=5 for each condition) was extracted from each well using Tri-Reagent (Invitrogen, Ireland) extraction, chloroform phase separation and isopropanol precipitation. cDNA was generated by reverse transcription of 0.5 µg of DNase treated total RNA using the random primer method (Invitrogen, Ireland).
Quantitative RT-PCR
RT-PCR TaqMan assay was used to quantitate the relative gene expression levels of the required genes and was performed on an ABI PRISM 7900HT Sequence Detection System (Applied Biosystems, Foster City, California, USA). Quantitative RT-PCR was carried out targeting COL6A3, CSPG2 and TGF-β1. The primers and probes for these targets were supplied as a preoptimised single tube primer Gene Expression Assay (Applied Biosystems). The endogenous control 18S rRNA was purchased from Applied Biosystems as a predeveloped assay reagent with the probe labelled with VIC at the 5′ end and TAMRA at the 3′ end to facilitate dual reporter assay and was examined in each sample. RT-PCR reactions were set up with Taqman Universal PCR Master Mix from Applied Biosystems.
Gene expression rates were compared using 18 s rRNA normalised threshold cycle number values (Ct). Subtracting the mean calibrator Ct from each individual sample yields ΔCt. The equation 2−ΔCt, was used to derive a fold difference in gene expression. When original Ct values for the target gene were above 40 cycles, it was designated as no detectable expression.
Statistical analyses
Gene expression data are presented as mean±SD. Data were analysed using one-way analysis of variance (ANOVA) with posthoc Bonferroni testing to account for comparison of the expression of individual genes at varying levels of strain in the presence or absence of verapamil. Statistical significance was set at p<0.05. Statistical analysis was performed using SPSS (IBM). Normality testing was carried out using the Kruskal–Wallis test. Calcium imaging data is presented as mean±SEM for a series of the indicated number of experiments. Statistical analysis was obtained using the Student's paired t test for analysis between two groups or one-way ANOVA and Tukey’s posthoc test for multiple analyses of more than two groups.
Results
LC cell [Ca2+]i changes induced by hypotonic strain
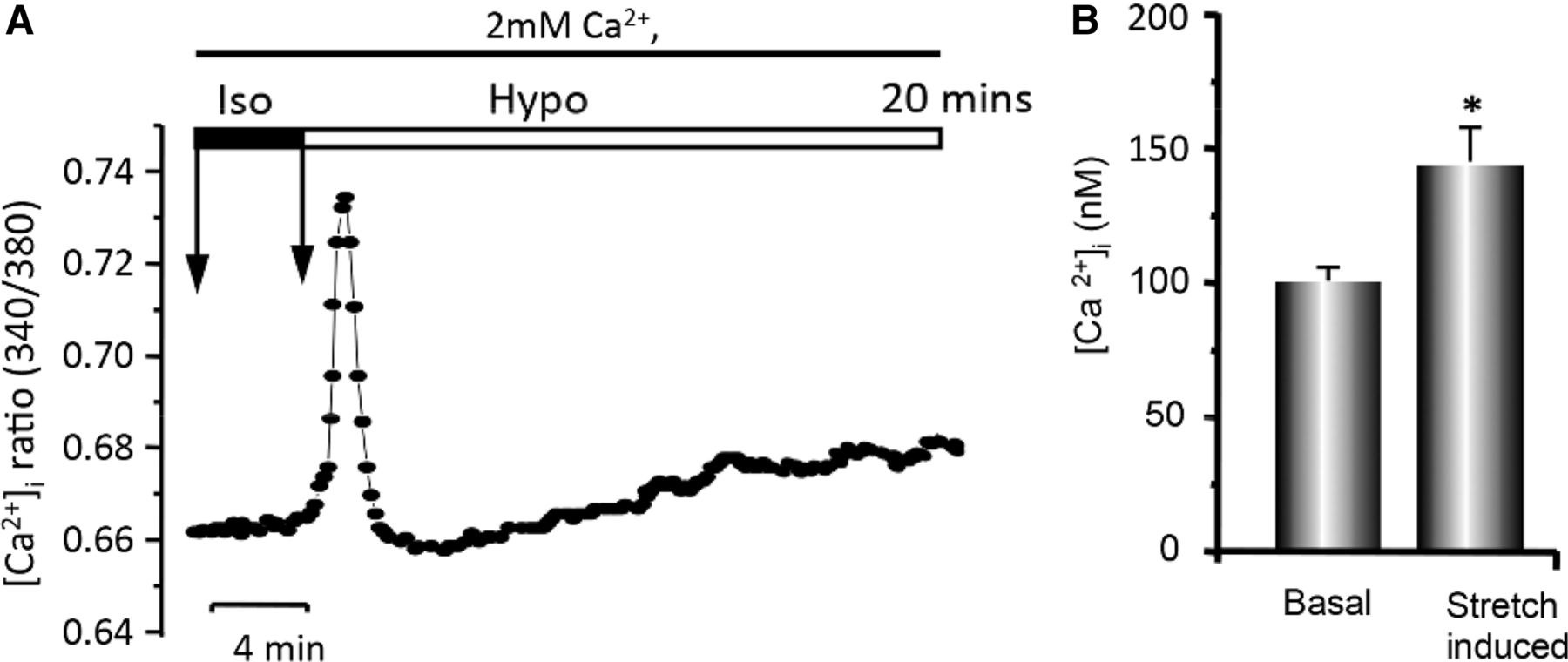
Calcium-imaging experiments were performed in fura-2/AM-loaded normal LC cells to evaluate [Ca2+]i changes under isotonic (physiological) and hypotonic stretch conditions in the presence of extracellular Ca2+ (2 mm). Hypotonic strain of LC cells significantly increased [Ca2+]i in a biphasic manner consisting primarily of a transient spike in [Ca2+]i reflecting the release of sequestered Ca2+ from internal stores followed by a secondary slight but sustained rise in [Ca2+]i due to Ca2+ entry from the extracellular space. Hypotonic stretch induced an increase in [Ca2+]i from 113 nm to 148 nm in this experiment (figure 2A). On average, hypotonic strain caused an increase in [Ca2+]i in LC cells, from 100±5.06 nm to 149±8.92 nm (p<0.05) (figure 2B).

Hypotonic stretch-induced changes in Ca2+ levels. (A) Representative time tracings of intracellular calcium, [Ca2+]i, ratio (340/380) in fura-2/AM-loaded lamina cribrosa (LC) cells in isotonic solution (Iso) followed by hypotonic stretch (Hypo) in the presence of 2 mM extracellular Ca2+. The changes in [Ca2+]i were taken from measuring peak values and expressing them as the change in fluorescence ratio from basal levels and determined for each experiment. The first arrow indicates the start of experiment and the second the timing of osmolarity change. (B) Mean transient peak increase of cytosolic Ca2+ in fura-2/AM-loaded LC cells. (*p<0.05, N=10 different experiments, n=69 cells for seven different LC donors. Note the significantly higher level of cytosolic [Ca2+]i in hypotonic stretch-induced conditions.)
The effect of verapamil on the hypotonic stretch-induced [Ca2+]i rise in LC cells
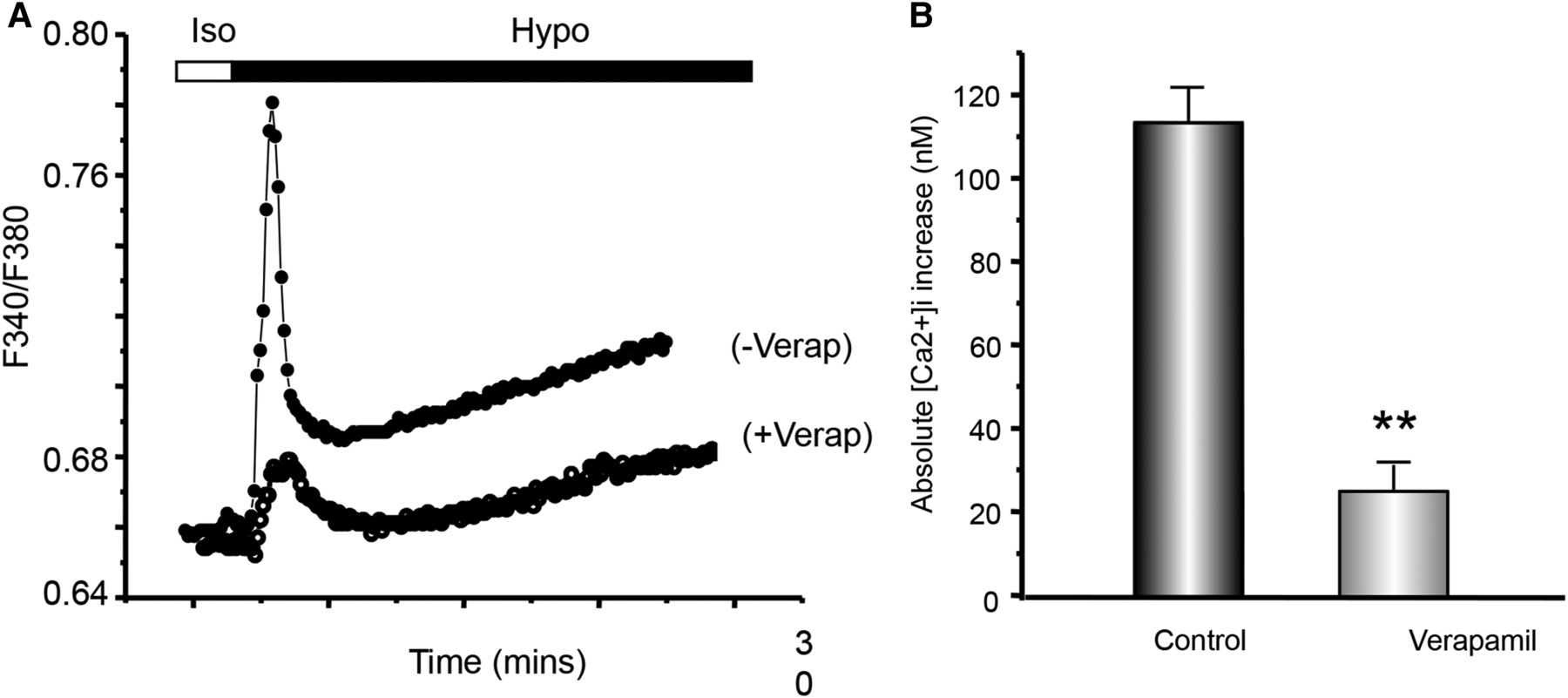
Calcium-imaging experiments were performed in fura-2/AM-loaded normal LC cells to evaluate the effects of verapamil on the hypotonic stretch-induced rise in [Ca2+]i. Verapamil significantly blocked the transient increase in the [Ca2+]i ratio induced by hypotonic solution (113.83±8.23 nm vs 25.34±6.44 nm, p<0.05) (figures 3A and B).

The effect of verapamil on the hypotonic stretch-induced intracellular calcium, [Ca2+]i, rise in lamina cribrosa (LC) cells. (A) Representative time tracings of [Ca2+]i ratio (F340/380) in fura-2-loaded normal LC cells. LC cells were first settled in isotonic (Iso) solution then stimulated by a hypotonic (Hypo) solution in the presence of 2 mM extracellular Ca2+. Cells were either pretreated (+verap) or non-treated control (−verap) with verapamil. Verapamil significantly blocked the transient increase in [Ca2+]i induced by the hypotonic solution. (B) Histogram comparing the mean [Ca2+]i increase (initial rapid, transient spike) under hypotonic conditions for LC cells in control non-treated versus those pretreated with verapamil. Note the significant difference in the [Ca2+]i rise under hypotonic conditions, 113.83±8.23 nM versus 25.34±6.44 nM; N=5 experiment, n=41 cells for three different control donors with no sign of glaucoma.
Positive identification of the α subunit of the L-type calcium channel
In order to ensure the presence of the L-type calcium channel for blockade, RT-PCR followed by agarose gel electrophoresis was carried out targeting the α1 subunit. The α1 subunit pore (∼190 kDa in molecular mass) is the primary subunit necessary for channel functioning in the high-voltage-gated calcium channels, and it consists of the characteristic four homologous I–IV domains containing six transmembrane α-helices each. Using agarose gel electrophoresis, the α subunit of the L-type calcium channel was identified in LC cells (figure 4).

Agarose gel electrophoresis for L-type calcium channel α subunit. The presence of L-type calcium channel α subunit was identified by PCR in lamina cribrosa (LC) cells (lanes 2–4). −ve control comprised of reaction without complimentary DNA was also identified (lane 5), (n=3).
The effect of verapamil on strain-induced ECM gene expression in LC cells from normal donors
Of the different experimental conditions outlined in figure 1, 15% strain provoked a statistically significant transcriptional response in all genes examined in normal LC cells (TGF-β1, COL6A3 and CSPG2) (figure 5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Normal lamina cribrosa (LC) cells were exposed to 0% or 15% strain for 24 h in the presence or absence of verapamil (10 mM). Total RNA was then harvested and the expression levels of three glaucoma related target genes were assessed by quantitative PCR. The graph shows the results for transforming growth factor-β 1 (TGF-β1), chondroitin sulfate proteoglycan 2 (CSPG2) and collagen 6A3 (COL6A3). 15% strain with verapamil showed a significant reduction in the expression of all three genes compared with 15% strain alone. Individual p values for each gene are given; n=3 per group. *p value=0.002, **p value=0.005, ***p value=0.021.
TGF-β1
Fifteen per cent strain induced a significant rise above baseline in TGF-β1 mRNA in normal donor LC cells (1.88±0.31-fold, p=0.002) (figure 5). With the addition of verapamil in the presence of 15% strain, the mRNA levels fell by 18% compared with the static baseline (0.82±0.14-fold, p=0.01). The decrease in transcriptional levels of TGF-β1 mRNA observed between the two 15% strain groups, in the presence or absence of verapamil, was statistically significant (p=0.001) (figure 5).
CSPG2
Fifteen per cent strain again induced a similar statistically significant rise above baseline levels of CSPG2 mRNA in human donor LC cells (4.64±1.64-fold, p=0.005) (figure 5). With the addition of verapamil in the presence of 15% strain, transcriptional levels fell by 2.64-fold (4.64±1.64 vs 2.0±0.211-fold, p=0.028). No statistically significant difference could be shown between 0% strain with or without verapamil and 15% strain with verapamil (figure 5).
COL6A3
Fifteen per cent strain induced an increase in COL6A3 mRNA levels (2.29±0.20-fold increase above baseline values, p=0.021) (figure 5). There was a statistically significant difference in COL6A3 mRNA levels between 15% strain in the absence of verapamil and 15% strain in the presence of verapamil (2.91±0.196-fold vs 1.118±0.417-fold, p=0.022) (figure 5).
Discussion
This is the first reported evidence that the ECM response of normal LC cells to significant mechanical strain can be modulated by Ca2+ antagonism, which may therefore be a possible future potential therapeutic target. This research supports the claim that LC cells can alter their surrounding ECM directly and indirectly (via growth factors) in a Ca2+ dependent manner in response to strain.
Studies by our group recently concluded that impaired Ca2+ extrusion in glaucoma LC cells compared with normal LC cells may be a target for future therapies.16 Furthermore, these findings compliment work demonstrating a higher level of [Ca2+]i in TM cells from patients with primary open-angle glaucoma (POAG).15 He et al concluded that the dysfunction in Ca2+ regulation (possibly secondary to mitochondrial dysfunction) by these cells may contribute to the failure of the TM to control IOP.15
In our experiments, we showed that hypotonic LC cell membrane stretch induced a transient peak followed by a gradual sustained increase in [Ca2+]i levels. Verapamil significantly blocked the transient increase in [Ca22+]i induced by the hypotonic solution. Ideally, we would have liked to examine the calcium transients induced by mechanical strain in the Flexercell system. However, since the LC cells are grown on flexible opaque membranes, it is not possible to conduct calcium-imaging experiments using the Flexercell system.
In an animal model of cardiac hypertrophy, Gao et al suggested that Ca2+ influx through L-type calcium channels is the primary source of Ca2+ to activate the nuclear factor of activated T-cells signalling pathway in cardiac myocytes.25 They concluded that while transient receptor potential (TRP) channels cause cardiac hypertrophy, they appear to do so through a mechanism involving activation of Ca2+ entry via L-type calcium channels. Extrapolating from the cardiac myocyte model, the ability to block this initiator of ECM remodelling could also signify a potential future therapeutic target in LC cells.
Previous work by our lab has examined the response of LC cells to graded mechanical strain and showed that a threshold level of strain appears to exist, after which the induction of ECM components begins (above 12% strain).22 We also demonstrated that by subjecting LC cells to 20% strain ,we induced an upregulation of COL6A3, CSPG2 and TGF-β1.22 This increase in transcription levels of ECM related genes was mimicked with an increase in protein levels, specifically TGF-β1 after 24 h. Similarly, in our current study, we have induced an upregulation in ECM related genes with 15% strain, which substantiates previous work by our lab.22
Verapamil inhibited the strain induction of TGF-β1, CSPG2 and COL6A3 mRNA gene expression. TGF-β1 has been shown to be induced by strain, and it also plays a critical role in the induction of ECM gene expression in LC cells.19 TGF-β signalling is mediated through a number of cellular pathways, some involving Ca2+ signalling,26 which has been implicated in POAG pathology. Ryu et al27 hypothesised that elevated [Ca2+]i and subsequent phosphokinase C activation resulted in an increased expression of TGF-β1 and its receptor in human mesenchymal stem cells. A similar transcriptional picture to that seen in glaucoma LC cells is observed when LC cells are stimulated by TGF-β1 alone.28
Modulation of TGF-β's strain-induced ECM response has been achieved in other cell types using verapamil.29 Xu et al30 were able to modulate the TGF-β1 fibrotic response of hepatic stellate cells using verapamil. Recent work examining the effect of verapamil on the mechanical strain-induced responses of bone marrow mesenchymal stem cells has shown that verapamil partially negated the strain-induced expression of c-fos, c-jun and c-myc.31 Recent work in cardiac myocytes has shown that verapamil induces a calcineurin-nuclear factor Y signalling pathway that can alter cardiac gene transcription and apoptosis.32 Given the results of our work, it is possible that a similar relationship involving [Ca2+]i, TGF-β and altered ECM gene expression exists in LC cells.
The use of calcium channel blockers in the treatment of glaucoma has been proposed for a number of decades. Calcium channel blockers have been proposed to lower IOP,33 increase ocular blood flow17 and be neuroprotective34 in glaucoma. Santafe et al35 carried out a number of animal studies that showed the IOP lowering properties of calcium channel blockers. Topical verapamil has also been shown to reduce IOP in normal humans36 as well as in humans with ocular hypertension.33 Recently, a number of review articles have called for randomised control trials to review the future therapeutic role of calcium channel blockers in glaucoma.34 ,37
This is the first study to show that calcium channel blockers have the ability to modulate the strain-induced ECM response of LC cells. Our current study provides intriguing evidence of a novel mechanotransduction pathway linking strain, cation channel functions and the induction of LC cell ECM gene transcription.
References
Footnotes
BQ and MI contributed equally.
Funding With generous support from a National Glaucoma Research (The American Health Assistance Foundation) grant and the Health Research Board (Ireland).
Competing interests None.
Patient consent Obtained.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- At a glance