Article Text

Abstract
Purpose Presence of SARS-CoV-2 RNA in human retinal biopsies (RBs) was previously reported by us. In this consecutive study, we analysed RB and optic nerve biopsies (ONBs) in deceased patients with confirmed COVID-19 assessing viral RNA load, possible virus replication and infectivity.
Patients and methods In this case series, 14 eyes of 14 deceased patients with COVID-19 were enucleated during autopsy. RB and ONB were subjected to molecular detection of viral RNA, virus cultivation and immunohistochemistry. SARS-CoV-2 RNA loads were compared with RNA loads in the respective throat swabs, vitreous humour and blood samples.
Results SARS-CoV-2 RNA was detected in 7/14 RBs and in 10/13 ONBs. While virus isolation failed and immunohistochemistry of SARS-CoV-2 spike protein was negative, subgenomic RNA (sgRNA) was detectable (40% RB; 60% ONB).
Conclusion SARS-CoV-2 RNA is detectable in RB and ONB of patients with COVID-19. Presence of sgRNA could point to a SARS-CoV-2 infection of neuronal tissue, but as virus isolation failed and immunohistochemistry of SARS-CoV-2 spike protein was negative, an active infection seems unlikely.
- COVID-19
- optic nerve
- retina
- microbiology
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
SARS-CoV-2, SARS-CoV and Middle East respiratory syndrome coronavirus (MERS-CoV) belong to the genus Betacoronavirus within the large family of Coronaviridae, share a similar structure and account for respiratory diseases with high mortality.1 2 Both SARS-CoV and SARS-CoV-2 enter the cell through binding the ACE2, which is expressed in different human structures including the brain3 and retina.4 SARS-CoV and MERS-CoV are known to cause neurological damage.5 Many studies also suggest a central nervous system (CNS) and peripheral nervous system involvement of SARS-CoV-2 induced COVID-19.2 3 5 The major route of CNS infection remains unclear. Suggested are among others haematogenic spread and spread via the olfactory bulb.3
In a study from Wuhan, China, 36.4% of COVID-19 infected patients showed neurological manifestations.6 Visual impairment occurred in 3 out of 214 (1.4%) of the patients. So far there is limited evidence supporting direct infection of the optic nerve.5 In a case report of a patient with COVID-19 developing diplopia, MRI showed enhancement of the optic nerve sheath and posterior Tenon capsules that could reflect leptomeningeal invasion.7
In a pilot study, we detected SARS-CoV-2 nucleic acid in human retina8 but we were unable to state where in the retina the virus might be located and if it replicates. In a clinical case series, hyper-reflective lesions were found in optical coherence tomography in 12 patients at the level of ganglion cell layer and inner plexiform layer of the retina.9 However, these findings have to be interpreted with great caution as this hyper-reflective lesions might just have been sections through normal inner retinal vessel as other authors have argued.10
Therefore, retinal biopsies (RBs), optic nerve biopsies (ONBs), vitreous humour (VH), throat swabs and blood serum from 14 deceased patients with COVID-19 were analysed to detect and quantify SARS-CoV-2 RNA by real-time reverse transcriptase PCR (RT-PCR). To understand infectivity, virus isolation was attempted. Immunohistochemistry on SARS-CoV-2 spike protein from RB and ONB was performed to further characterise a possible infection of the posterior segment of the eye with SARS-CoV-2.
Methods
Study design
Between 20 March and 14 May 2020, 170 autopsies of deceased patients with COVID-19 were performed at the Institute of Legal Medicine at the University Medical Center Hamburg-Eppendorf. Autopsies were arranged according to §25 of the German Infection Protection Law. The study complied with the tenets of the Declaration of Helsinki. Included were patients with a positive throat swab on SARS-CoV-2 RNA during lifetime.
All deceased patients were screened for SARS-CoV-2 RNA using a throat swab followed by immediate RT-PCR. Retinal biopsies, ONB, VH, throat swab and venous blood were taken for further analysis. Clinical records were checked for pre-existing medical conditions and antemortem diagnostic findings.
Retina preparation and VH sample
Until autopsy, the bodies were stored at 4°C. For RB, the eyes were enucleated and dissected at the equator of the bulb. Consequently, the vitreous automatically detached from the retina as the anterior part of the eye was removed. A native (fresh, unfixed tissue) sample of the VH was taken. A peripheral retinal detachment was induced before the native RB was taken to receive a pure RB. The centre of the retina was left untouched and adjacent for immunohistochemistry. For immunohistochemistry, the dissected bulb was then fixed in buffered 4% formaldehyde.
Optic nerve preparation
The optic nerve was collected after opening the cranial cavity during autopsy on the level of the optic chiasm. Two samples were taken: one native and the other fixed in buffered 4% formaldehyde. The native samples were directly transferred to the in-house Institute of Microbiology, Virology and Hygiene for further analysis. The formaldehyde fixed samples were transferred to the in-house Institute of Neuropathology for histology and immunohistochemistry.
Molecular diagnostic
Native tissue samples were obtained during autopsy, transferred to microtubes containing ceramic beads (Precellys lysis kit) and ground in 1 mL of sterile and RNAse free H2O using an appropriate homogeniser (Precellys 24, Bertin, USA). For detection of SARS-CoV-2 in tissue samples, automated nucleic acid extraction (MagNA Pure 96 (Roche)) was performed, and subsequent SARS-CoV-2 RT-PCR was performed on a Light Cycler 480 II instrument (Roche, USA) as described previously,11 using the one-step RNA control kit (Roche). As a marker for nucleic acid extraction quality and to allow for normalisation of viral RNA load in tissue samples, single copy human ß-globin gene was quantified by PCR (Life Technologies, Thermo Fischer, USA). Detection of SARS-CoV-2 RNA in cell culture supernatant was performed as described previously.12 Quantitative RT-PCR for N-gene and subgenomic RNA (sgRNA-N gene) in homogenised tissue samples was performed as previously published13 with slight adaptations: N-gene sgRNA gene was quantified combing a 5-UTR region primer (leader sequence, sgLeadSARSCoV2-F; 5-CGATCTCTTGTAGATCTGTTCTC-3′13 with a reverse primer (2019-nCoV_N1-R, 5′-TCT GGT TAC TGC CAG TTG AAT CTG-3′ and probe (2019-nCoV_N1-P, 5′-FAM-ACC CCG CAT TAC GTT TGG TGG ACC-BHQ1-3′ binding to the nucleotide gene. SARS-CoV-2 (isolate HH-114) infected Vero cells (ATCC CRL-1586) served as a control.
Cell culture and virus isolation
Vero cells (ATCC CRL-1586) were cultivated in Dulbecco's Modified Eagle Medium (DMEM) containing 10% fetal calf serum, 1% penicillin/streptomycin, 1% L-glutamine, (200 mM), 1% sodium pyruvate, 1% non-essential amino acids (all Gibco/Thermo Fisher, Waltham, USA) under standard culture conditions. For virus isolation attempts, cells were seeded to 80%–90% confluency into 24-well plates (TPP, Switzerland). For infection, 250 µL of the homogenised tissue solution was added to each well. Adsorption was performed for 1 hour at 37°C. Subsequently, cells were washed once with phosphate buffered saline and 1 mL of fresh cell culture medium was added. Cells were monitored daily for cytopathic effect. Absence of virus growth was confirmed by quantitative RT-PCR.12
Immunohistochemistry
Formalin-fixed paraffin embedded tissue (FFPE) samples from the retina and optic nerve were processed and stained with H&E following standard laboratory procedures. For immunohistochemistry of SARS-CoV-2 spike protein, antibody (1:300, GeneTex; #GTX632604; clone 1A9, Irvine, USA) was validated on SARS-CoV-2 infected (Hamburg isolate14) and non-infected Vero cells that were processed to FFPE blocks. Slides were examined by experienced morphologists (MG and SK), and selected slides were electronically scanned at high magnification (×40) as high-resolution images (1900×1200 pixels) using a NanoZoomer 2.0‐HT (Hamamatsu Ltd, Hamamatsu, Japan).
Results
Patient cohort
Thirteen of the 14 analysed patients (93.86 %) died of COVID-19. One patient (7.14 %) died of a haemorrhagic shock most likely unrelated to his COVID-19 infection and was defined as non-COVID-19 death. The average age of patients was 70 years±19.3 years. The average postmortem interval (PMI) was 4±4 days. Details of patient characteristics and risk factors for severe chronic condition are given in table 1. No specific ocular symptoms in relation to COVID-19 were listed in any medical record.
Patient characteristics and risk factors for severe chronic condition
SARS-CoV-2 genomic and subgenomic RNA in the retina and optic nerve
SARS-CoV-2 RNA was detected in 7/14 RB (50 %), 10/13 ONBs (76.92 %) and 8/14 (53.85 %) VH tissue samples (figure 1A,B). Postmortem SARS-CoV-2 viraemia was detected in 6/11 (54.55%) (figure 1A). In samples that tested positive for genomic SARS-CoV-2 RNA, sgRNA was detected in 2/5 (40%) RBs and 6/10 (60%) ONBs (figure 1C). Virus isolation failed in all RB (14/14), ONB (13/13) and VH (14/14) samples. For all results, see online supplemental etable1.
Supplemental material

Detection of SARS-CoV-2 RNAs. SARS-CoV-2 RNA loads in eye samples, blood and throat swabs are displayed for each patient (patient number on the x-axis) (A); blank cells reflect SARS-CoV-2 RT-PCR negative samples, and X indicates samples that were not available for analysis. For eye samples, virus-to-cell ratio (B) and levels of sgRNA (C) are illustrated. Mean values are given by horizontal bars. Negative samples are shown below the horizontal bar marked ‘neg’. SARS-CoV-2 infected Vero cells served as positive control, while cell culture supernatants of the respective cultures served as negative control. sgRNA, subgenomic RNA.
Histology
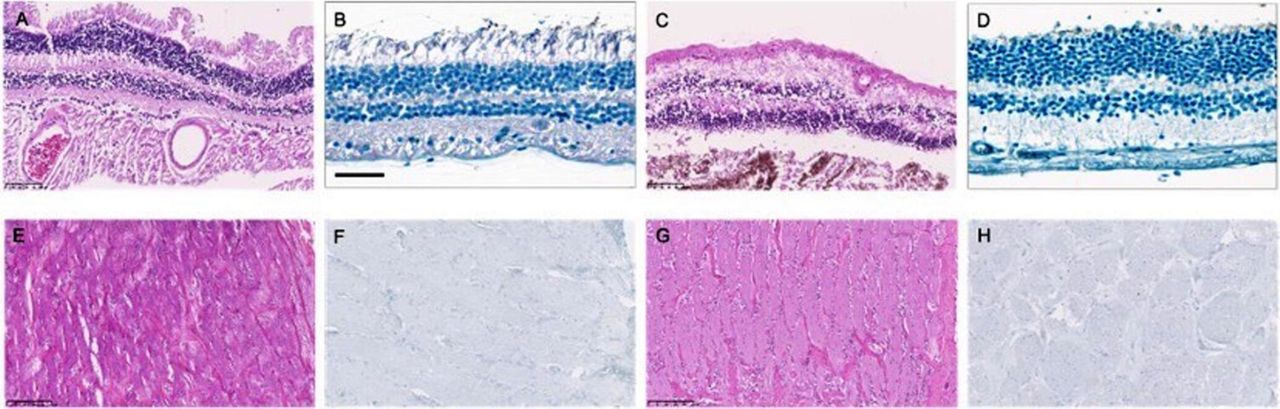
Representative pictures of one SARS-CoV-2 PCR negative and positive sample of RB and ONB are displayed in figure 2. H&E staining of the RB reveal a preserved retinal structure in SARS-CoV-2 PCR negative and positive samples (see figure 2A,C). No obvious differences were seen between PCR negative and positive ONB samples (figure 2E,G).
{kind=link}
{kind=link}
Histology and immunohistochemistry: retina and optic nerve. Retinal sections of COVID-19 deceased patients are displayed in figure panels A–D, sections of the optic nerve respectively in figure panels E–H. (A and B) PCR-negative retina. (A) H&E staining, 20× magnification; (B) SARS-CoV-2 spike protein. (C and D) PCR-positive retina. (C) (H&E), 20× magnification; (D) SARS-CoV-2 spike protein. (E and F) PCR-negative optic nerve. (E) H&E staining, 20× magnification; (F) SARS-CoV-2 spike protein. (G and H) PCR-positive optic nerve.
Immunohistochemistry: SARS-CoV-2 spike protein
SARS-CoV-2 spike protein positive cells could not be detected by immunohistochemistry in RB and ONB regardless of a positive or negative SARS-CoV-2 PCR (see figure 2B,D,F,H).
Discussion
Recent studies have shown that SARS-CoV-2 uses ACE2 as receptor for entry.15 16 In humans, ACE2 was detected on the ocular surface,17 in the aqueous humour,18 the retina4 and the brain.3 ACE2 activity was found in the ciliary body, vitreous body and retina of porcine ocular tissue.19 Neurological manifestations occur in approximately 1/3 (36.4%) of COVID-19 infected patients.6 Visual impairment was reported to be rare in patients with COVID-19 (3 out of 214 patients),6 but in our study, in 10 out of 13 ONBs and 7 out of 14 RBs, viral RNA could be detected by RT-PCR in postmortem tissue samples. However, the lack of visual impairment does not directly imply the absence of active ocular involvement in patients with COVID-19. Preserved visual acuity is not uncommon in smouldering inflammatory ocular diseases like tuberculosis20 or sarcoidosis21 associated posterior uveitis.
In a previous pilot study,8 we were able to detect SARS-CoV-2 nucleic acid in human retina, but due to the lack of blood and VH samples, we could not rule out a possible contamination of the RB. In previous studies, a positive RT-PCR result on SARS-CoV-2 was found in 1% (3 out 307),22 8% (1 out 12)23 and 27% (19/71)24 of blood samples of patients with COVID-19. In our study, the rate of positive blood samples was 54.6% (6/11) compared with a rate of SARS-CoV-2 RNA of 50% (7/14) in RB and 76.9% (10/13) in ONB. The high rate of postmortem viraemia could be due to the fact that preferably patients with a low CT-value in the throat swab during lifetime (eg, high viral loads) were analysed. Furthermore, high plasma viral load is associated with worse disease severity and higher mortality.24 Interestingly, analyses identified three patients (patients #4, #8 and #12) without detectable viraemia and three patients (patients #4, #5 and #8) with negative postmortem throat swabs but ONB and/or RB tested positive in RT-PCR on SARS-CoV-2. Throat swabs turn negative depending on the days after onset of symptoms.13 Five days after the onset of symptoms, viral load decreases, and detection rate is at 39.93%.13 Therefore, these negative postmortem throat swabs do not challenge the positive results in RB and/or ONB.
Vice versa, one patient (patient #13) presented with a positive blood sample but negative result in RB, ONB and vitreous body. Consequently, no direct correlation seems to exist between viraemia and a SARS-CoV-2 positive RB or ONB. In patient #3, the viral RNA load in the RB and ONB was higher than in the blood sample. In patient #1, the viral RNA load in RB and ONB was still roughly half of the level detected in blood sample. Five (patients #2, #4, #5, #6 and #11) out of 14 patients died of either a haemorrhagic shock or sepsis (see table 1) where vascular and cellular integrity is compromised, which could possibly lead to contamination. However, because also non-septic patients tested positive for SARS-CoV-2 RNA in RB and ONB, we consider the risk for contamination to be low.
Up to date, there is no proof of SARS-CoV-2 presence in the VH. List et al 25 reported on the absence of SARS-CoV-2 in the VH. However, in that study, no data were reported on the viral RNA load in blood samples. The high rate of viraemia in our study and thus higher viral loads could be the reason why SARS-CoV-2 RNA was detected. Furthermore, the presence of ACE2 receptors support our findings of positive SARS-CoV-2 RT-PCR results in VH. A spillover on the retina with SARS-CoV-2 through VH is possible but unlikely. To eliminate this confounder, we removed the vitreous in total before obtaining the RB. The ONBs were obtained through a cranial access leaving the intrabulbar part of the optic nerve intact. Therefore, a contamination of the ONB via VH is also unlikely, which is supported by 10 out of 13 ONB samples testing positive in our study.
In summary, our data on blood and VH make contamination as source for the detection of SARS-CoV-2 RNA in human retina and optic nerve unlikely.
Until today, the major route of CNS infection remains unclear, but in discussion are among others haematogenic spread and spread via the olfactory bulb.3 Retina, as a part of the CNS, could possibly be infected through haematogenic spread and also through nerve fibres via the optic nerve. The viral RNA loads of the ONB are higher than in RB. RBs were only positive if the ONB was positive as well but not vice versa. This could be a possible hint on antegrade neural spread of the virus from the optic nerve into the retina.
In addition, in those samples tested positive on SARS-CoV-2 RNA, sgRNA was detected in 40% and 60% of RB and ONB samples, respectively. While sgRNAs certainly are a replication intermediate and are not present in viral particles, we cannot conclude that detection reflects actually viral replication in these tissues. Subgenomic RNA might also be based on cells temporarily present in the analysed tissue (eg, migrating immune cells) or RNA residues of dead cells from other tissues.26
In contrast to the positive results regarding genomic and sgRNA, no infectious virus could be recovered from any investigated eye tissue. Virus isolation in cell culture in general is an insensitive method. Success of virus isolation strongly depends on viral loads and the time after COVID-19 diagnosis as shown by previous studies.27 As most of the eye samples had rather low RNA levels when compared with oropharyngeal swabs or lung samples (that are usually used for virus isolation attempts), virus isolation failure for those samples was expected. In other eye samples, especially in the optic nerve, a higher viral load was detected. This amount of viral RNA measured suggests that virus isolation in cell culture should be possible. However, variable PMIs might interfere with successful virus isolation. Also, the time after infection plays a significant role for infectivity and the success of virus isolation, as most individuals will produce neutralising antibodies and thus RNA titres do not correlate with infectivity in those patients.13 28 29 These findings most likely indicate that there is no viable virus in the tissue, but due to other possible reasons for failure of cell culture growth, it cannot fully be concluded on non-viable virus in the tissue.
In line with the negative results of virus cultivation, SARS-CoV-2 spike protein visualisation in RB and ONB by immunohistochemistry was also not successful. This diagnostic tool though is also by far less sensitive than a RT-PCR on viral RNA.
Limitations to this study are the small sample size and that only a single section was taken for investigation. Small sample sizes always pose the risk of overestimating the conclusion; therefore, these are summed up conservatively. Retrieving multiple biopsies could have helped to rule out contamination and to support positive results. As the dissected bulb was saved for immunohistochemistry, we tried to keep it as intact as possible. Therefore, only a single RB was taken.
In conclusion, we confirm our previous result that SARS-CoV-2 RNA can be detected in the retina of patients with COVID-19. Additionally, we show that SARS-CoV-2 nucleic acid can also be found in VH and ONB. SgRNA was detectable in almost half of RB and ONB samples; however, sgRNA is not a definite indicator of active replication. Virus isolation and immunohistochemistry on SARS-CoV-2 spike protein failed in all samples. As a previous case series has shown, there is a clinical impact of SARS-CoV-2 on retinal structures,9 but visual impairment is rare.5 Moreover, active infection within these ocular tissues seems unlikely. Further research is necessary to investigate a possible ophthalmological impact in a larger cohort of patients with COVID-19.
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information.
Ethics statements
Patient consent for publication
Ethics approval
Institutional Review Board approval from the independent ethics committee of the Hamburg University was obtained (protocol-no PV7311).
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
MC and AF contributed equally.
Funding This study was funded by DEFEAT pandemics: work was partially supported by a grant of the research consortium DEFEAT PANDEMics – National Network University Medicine (NUM) funded by the Federal Ministry of Education and Research (BMBF) (grant number: 01K×2021).
Competing interests MS reported receiving grants from Novartis, IDxDR and Boehringer Ingelheim; and personal fees from Bayer, Oxurion, Roche, Allergan, Alcon, Neurogene and GSK outside the submitted work.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.
Linked Articles
- Highlights from this issue
- Correction