Article Text

Abstract
BACKGROUND/AIMS Mechanical forces are thought to induce cellular responses through activation of signalling pathways. Cells within the intraocular environment are exposed to constant changes in the levels of intraocular pressure. In this study, an attempt was made to determine the acute effects of elevated hydrostatic pressure on different intraocular cells grown in culture.
METHODS Different cell lines derived from ocular tissues including non-pigmented and pigmented ciliary epithelium, trabecular meshwork, retina, and lamina cribrosa were incubated in a pressurised chamber at 50 mm Hg in a culture incubator at 37°C for up to 6 hours. Control cells were incubated at atmospheric pressure. The viability of the cells was examined using their intracellular esterase activity. The morphology and cytoskeleton of the cells were investigated using microscopy and phalloidin staining. Adenylyl cyclase activity was assessed by measuring the conversion of [3H]-cAMP from [3H]-ATP in response to elevated hydrostatic pressure for 1–6 hours. In addition, at the end of incubation period under elevated hydrostatic pressure the recovery of adenylyl cyclase activity to control levels was examined.
RESULTS Cell viability did not change following exposure to elevated hydrostatic pressure for 6 hours. Cells subjected to elevated hydrostatic pressure demonstrated morphological differences characterised by a more rounded shape and a redistribution of actin stress fibres that was most prominent in lamina cribrosa astrocytes. A time dependent increase in basal adenylyl cyclase activity, and a decrease in maximum forskolin stimulated activity were observed in all cell lines following exposure to elevated hydrostatic pressure.
CONCLUSION These observations demonstrate that cell lines from different ocular tissues are sensitive to changes in external pressure in vitro. They exhibit morphological and cytoskeletal changes as well as significant alterations of intracellular adenylyl cyclase activity following exposure to acute and sustained levels of elevated hydrostatic pressure of up to 6 hours' duration.
- adenylyl cyclase
- astrocytes
- ciliary epithelium
- hydrostatic pressure
- retina
- trabecular meshwork
Statistics from Altmetric.com
In vivo, most living cells are exposed to a variety of biomechanical forces by virtue of their relation with the microenvironment. Intracellular mediator pathways by which cells respond to biomechanical deformation are widely explored in biology. It has been shown that increased production of cAMP, prostaglandin E2, or prostacyclin may mediate the effects of compressive pressure on cells in culture.1-3 Studies of different cells from the human body reveal the effects of elevated hydrostatic pressure on cell shape and function. For example, in endothelial cells exposed to elevated hydrostatic pressure, release of bFGF can be induced independently of cell injury or death and may account for subsequent morphological and proliferative responses.4Similarly, elevated hydrostatic pressure is an important stimulator of matrix synthesis in cartilage. This results in the increased expression of cytokines and heat shock proteins thought to be necessary for the repair of damaged or disorganised cytoskeleton.5 Regarding ocular tissues, it has been shown that human lamina cribrosa cells may react to pressure changes in their environment by modulating the production and secretion of extracellular matrix macromolecules such as seen for collagen type I.6
Elevated intraocular pressure has been proposed to be an important risk factor for the development of glaucomatous optic neuropathy7 8 and many ocular cells are exposed to constant variations of intraocular pressure, which generate mechanical forces affecting cell behaviour. However, individual cellular responses of ocular tissues to elevated pressure or to pressure related stress remain largely unstudied. In this study, we investigated the responses of different ocular cells to elevated hydrostatic pressure by examining the cell viability and morphology. We also studied whether cells derived from ocular tissues demonstrate biochemical alterations of basic intracellular enzyme pathways such as adenylyl cyclase following exposure to sustained elevation of hydrostatic pressure for several hours. We present evidence that acute and sustained elevated hydrostatic pressure can affect cell morphology as well as alter adenylyl cyclase activity in cell lines derived from several intraocular tissues including the ciliary epithelium, trabecular meshwork, retina, and lamina cribrosa.
Methods
CELL CULTURES
Non-pigmented and pigmented ciliary epithelial cells used in our experiments were clones derived from human (non-pigmented) or bovine (pigmented) cells transformed with simian virus 40 (SV-40) as previously described.9-11 Transformed human trabecular meshwork cells were provided as a gift by Dr L DeSantis (Alcon Laboratories, Fort Worth, TX, USA).12 Ciliary epithelial and trabecular cells were grown in Dulbecco's modified Eagle's medium (DMEM) with 5% fetal calf serum and 5% each of penicillin and streptomycin. An immortalised rat retinal cell line (E1A.NR3) was provided by Dr G M Seigel which contains cells expressing antigens specific for photoreceptors, bipolar cells, ganglion cells, and retinal glial cells was used.13 These cells were maintained in DMEM supplemented with 10% fetal bovine serum and 1% each of non-essential amino acids, l-glutamine, vitamins, and antibiotics. Third to fourth passages of human lamina cribrosa astrocytes were cultured in DMEM/F-12 supplemented with 10% fetal bovine serum, 5 ng/ml of human basic pituitary fibroblast growth factor (Biomedical Technologies, Stoughton, MA, USA), 5 ng/ml of human platelet derived growth factor-A chain (Sigma, St Louis, MO, USA), and PSFM (10 000 U/ml penicillin, 10 000 μg/ml streptomycin, and 25 μg/ml amphotericin B) as previously described.14 All tissue culture reagents were purchased from Gibco (Gaithesburg, MD, USA).
STUDY DESIGN
Cells plated in six well plates (Costar, Cambridge, MA, USA) at a density of 3–10 ×104 cells/well and grown to semiconfluence in 2–7 days were incubated with serum free medium overnight. A closed pressurised chamber (92% air, 8% carbon dioxide) equipped with a manometer, that can be placed in a tissue culture incubator, was used so that the cells in six well plates could be exposed to elevated hydrostatic pressure.6 Cells were placed in the chamber and the pressure was elevated to 50 mm Hg. The chamber was subsequently placed in a tissue culture incubator at 37°C. To examine the time course of cellular responses, the elevated pressure was maintained for 1, 3, or 6 hours. Control cells from identical passage of cell lines were simultaneously incubated in a tissue culture incubator at atmospheric pressure at 37°C. At the end of the incubation period, the cells were immediately subjected to experiments described below. In addition, some of the plates incubated under elevated hydrostatic pressure were transferred to normal atmospheric pressure condition for 3 or 24 hours in order to examine the recovery of cell responses.
We examined the viability and morphology of the cells incubated in the presence or absence of elevated hydrostatic pressure. Furthermore, we assayed basal and forskolin stimulated adenylyl cyclase activity in addition to somatostatin attenuation of forskolin stimulation in order to assess the effects of hydrostatic pressure on the activity of this signal transduction pathway. The viability of the cells were determined with the Live/Dead Kit (Molecular Probes, Eugene, OR, USA) which contains calcein and ethidium homodimer.15 It relies on the intracellular esterase activity within living cells to cleave calcein to form a green fluorescent membrane impermeable product. In dead cells ethidium can easily pass through the compromised plasma and nuclear membranes and attach to the DNA yielding red fluorescence. At least 750 cells were counted under 200× magnification in three randomly chosen areas using a fluorescence microscope equipped with fluorescein and rhodamine filters (Olympus, Tokyo, Japan). The number of cells was counted using a Coulter counter (Coulter, Hialeah, FL, USA) after trypsinisation of the cells. The viability of the cells was expressed as the average ratio of esterase (+) cells to the total number of cells counted, multiplied by 100.
In addition to phase contrast microscopy to examine cell shape in response to elevated hydrostatic pressure, we performed F-actin staining using phalloidin to assess the status of actin cytoskeleton. At the end of the incubation period in the presence or absence of elevated hydrostatic pressure, cells grown on coverslips were immediately fixed in acetone at −20°C for 20 minutes, blocked using 5% horse serum for 30 minutes, and washed in phosphate buffered solution (PBS) containing 1% bovine serum albumin. The coverslips were then incubated with 50 μl of Oregon Green 48-phalloidin (Molecular Probes) for 30 minutes. After washing with PBS, the coverslips were mounted in ProLong Antifade medium. Pressurised and control cells stained with phalloidin were observed and photographed with a Zeiss LSM 410 laser confocal microscope.
ADENYLYL CYCLASE ACTIVITY ASSAY
After the incubation in the presence or absence of elevated hydrostatic pressure for up to 6 hours, cells were washed three times with PBS. After aspiration of PBS, 1.75 ml of a solution containing 1 μCi adenine [2,8-3H] (Dupont, Boston, MA, USA) per well prepared in DMEM, 2 mM HEPES, and 50 μl of 10 mM 3-isobutylmethylxanthine (IBMX) (Sigma) were added to each well. During this treatment period cells were incubated at 37°C with or without elevated hydrostatic pressure application for 20 minutes. At the end of the pre-incubation period for 3H-adenine labelling, cells were treated with DMEM, 100 μM forskolin (Calbiochem, La Jolla, CA, USA) solution, or 100 μM forskolin plus 2 μM somatostatin (Calbiochem) for 20 minutes at 37°C with or without elevated hydrostatic pressure application. At the end of the 20 minutes of drug incubation period, 4% ice cold trichloroacetic acid was added to each well to terminate the reaction. After incubation on ice for 30 minutes, the supernatant was collected from each well, spun for 3 minutes in a Beckman microfuge, and used to assay the conversion of [3H]-cAMP from [3H]-ATP following chromatography over Dowex ion exchange resin (Bio-Rad, Richmond, CA, USA) columns as previously described.16 The percentage of conversion of [3H]-ATP to [3H]-cAMP was used to express the adenylyl cyclase activity. The experiments described above were repeated three times for each cell line and means (SD) of the three experiments were presented.
Results
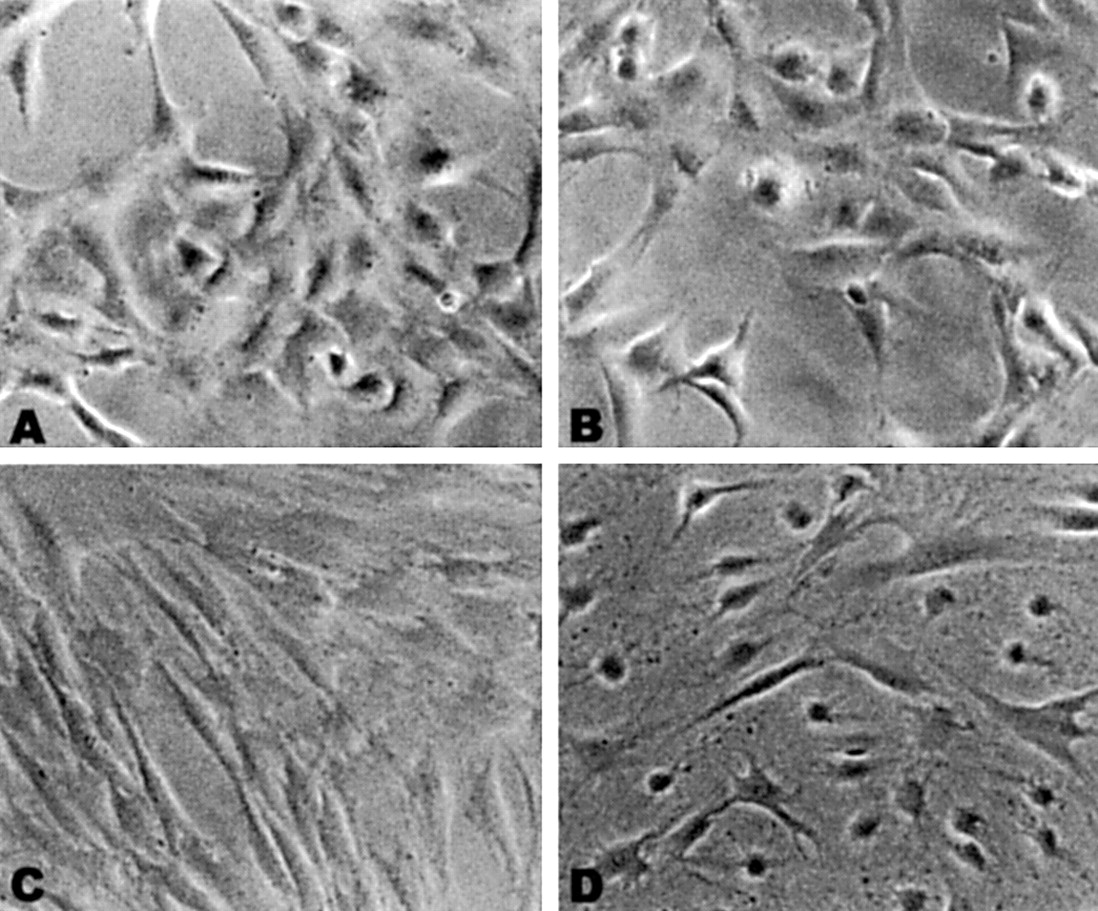
Exposure to elevated hydrostatic pressure maintained at 50 mm Hg for up to 6 hours caused no detectable change in cell viability as assessed by intracellular esterase activity (Table 1). However, the cell morphology was changed with exposure to elevated hydrostatic pressure in all cell lines studied. These changes were characterised by a more rounded shape of the cells that was most prominent in lamina cribrosa astrocytes and trabecular meshwork cells (Fig 1).
Viability of the cells

Morphological changes of cells exposed to sustained elevated hydrostatic pressure. Phase contrast micrographs of living trabecular meshwork cells (A, B) and lamina cribrosa astrocytes (C, D). Note in (B) and (D) that cells exposed to elevated hydrostatic pressure to 50 mm Hg for 6 hours adopt a rounder shape when compared with control cells shown in (A) and (C) (original magnification ×20).
To visualise changes in the distribution and organisation of the F-actin cytoskeleton, cells were stained with fluorescent labelled phalloidin, which binds to actin filaments. Human trabecular meshwork cells showed a diffuse and fine filamentous staining in the cytoplasm. When cells were exposed to elevated hydrostatic pressure, redistribution of the actin stress fibres, retraction of processes, and an enhanced polygonal cell shape were observed (Figs 2A and B). Human laminar astrocytes exhibited distinct and prominent stress fibres extending across the cell cytoplasm in control cells. In lamina cribrosa astrocytes exposed to elevated hydrostatic pressure, F-actin staining revealed displacement of the stress fibres to the periphery of the cell cytoplasm in most cells (Figs 2C and D).

Effects of elevated hydrostatic pressure on F-actin cytoskeleton. (A) Human trabecular meshwork cells incubated at atmospheric pressure. (B) Human trabecular meshwork cells exposed to elevated hydrostatic pressure. Although control cells shown in (A) exhibit short processes and a diffuse filamentous staining, cells exposed to elevated hydrostatic pressure shown in (B) exhibited redistribution of actin fibres and retraction of processes. (C) Lamina cribrosa astrocytes incubated at atmospheric pressure. (D) Lamina cribrosa astrocytes incubated at elevated hydrostatic pressure for 6 hours. Note in (D) the displacement of phalloidin stained actin stress fibres to the periphery of the cell. Control cells shown in (C) are elongated and actin stress fibres are distributed throughout the cytoplasm.
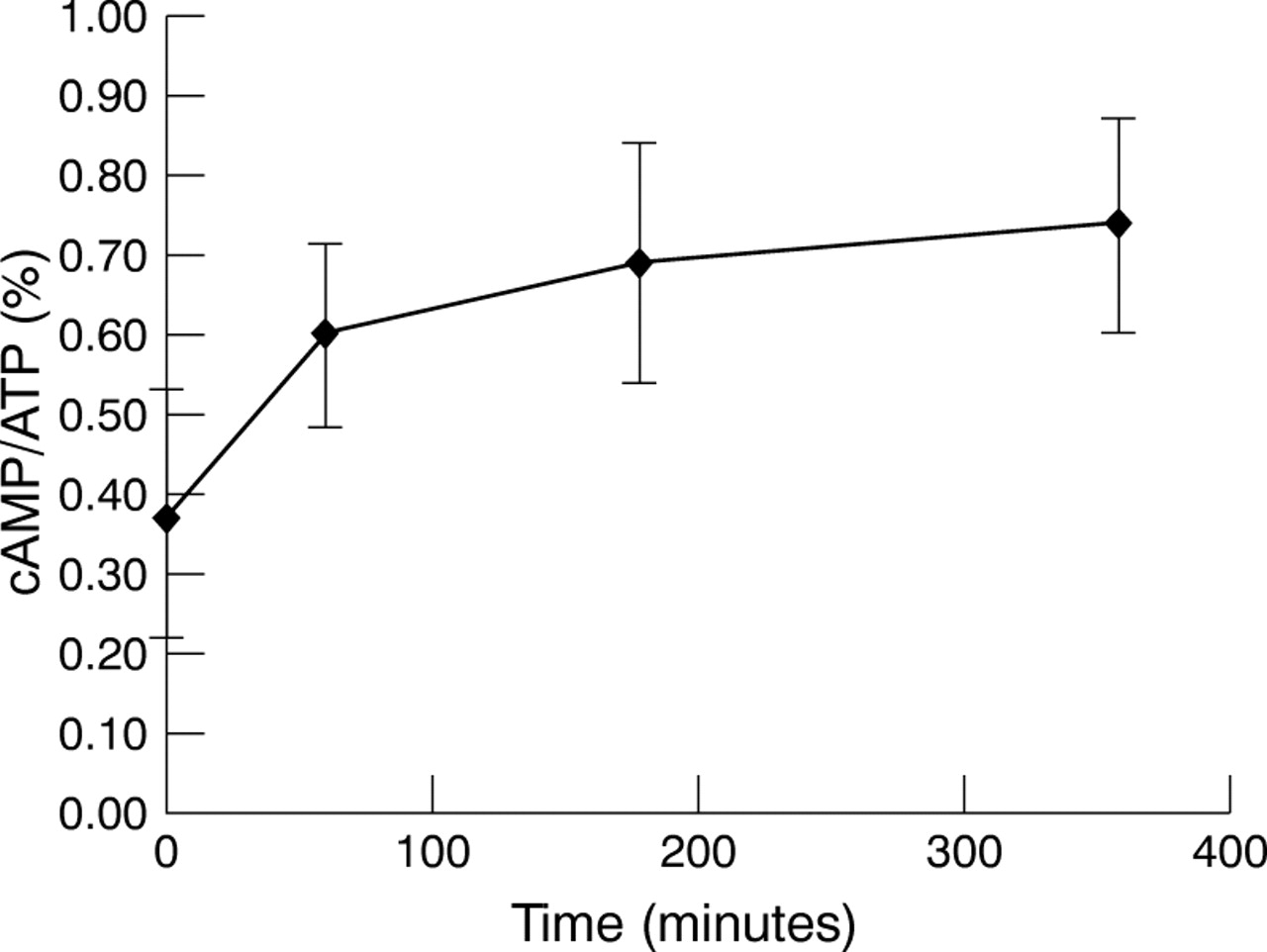
Basal adenylyl cyclase activity, as assessed by the percentage of [3H]-cAMP converted from [3H]-ATP, was comparable in all cell lines studied and ranged between 0.2% and 0.4%. Incubation of the cells under elevated hydrostatic pressure resulted in a time dependent increase in the adenylyl cyclase activity (Fig 3). The increase in the basal adenylyl cyclase activity of all cell lines after exposure to elevated hydrostatic pressure for 6 hours is shown in Figure 4. There was no statistical difference among the cell lines in either their control basal adenylyl cyclase activity or the increased cyclase activity following exposure to elevated hydrostatic pressure (multiple comparison test17, p >0.05). If the cells incubated under elevated hydrostatic pressure for 6 hours were then transferred to normal pressure condition, adenylyl cyclase activity decreased to their initial levels at a rate that was dependent on the time of exposure to elevated hydrostatic pressure. When cells were incubated under elevated hydrostatic pressure for 1 hour, a return to basal levels were observed within 3 hours. However, the recovery period was longer if the cells were incubated under elevated hydrostatic pressure for 6 hours; cells returned to approximately 70% of their baseline values after 3 hours, with full recovery after 24 hours (data not shown).

Time course of changes seen in the basal adenylyl cyclase activity of lamina cribrosa astrocytes exposed to elevated hydrostatic pressure up to 6 hours.

Percentage of increased basal adenylyl cyclase activity in different cells from ocular tissues with elevated hydrostatic pressure. NPE = non-pigmented ciliary epithelium; PE = pigmented ciliary epithelium; TM = trabecular meshwork cells; A = lamina cribrosa astrocytes; R = retina cells.
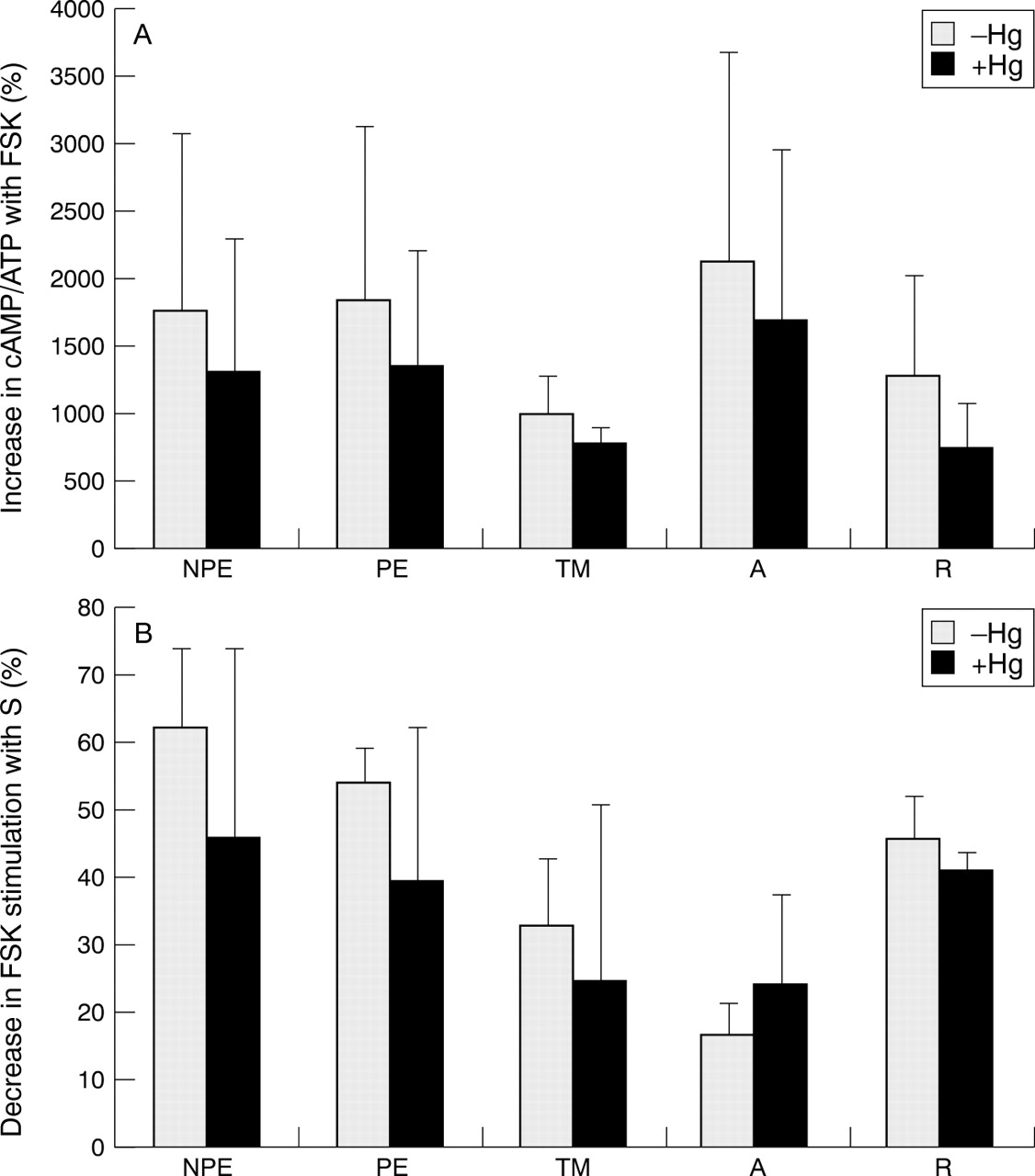
In order to assess the capability of cells to maximally activate adenylyl cyclase, individual cell lines were treated with 100 μM forskolin in the presence or absence of elevated hydrostatic pressure. Forskolin stimulation of adenylyl cyclase activity was decreased similarly (∼15–20%) in all cell lines studied (multiple comparison test, p >0.05) following 6 hours' exposure to elevated hydrostatic pressure (Fig 5A). We also utilised somatostatin to study the ability of cells to inhibit adenylyl cyclase activity following their maximal stimulation with forskolin. Two pertinent findings were noted. Firstly, somatostatin attenuation of forskolin stimulated adenylyl cyclase activity was lower in lamina cribrosa astrocytes compared with other cell lines either in the presence or absence of elevated hydrostatic pressure (multiple comparison test, p <0.01). Secondly, although the difference between the response of astrocytes to somatostatin treatment in the presence or absence of elevated hydrostatic pressure did not reach statistically significant levels, astrocytes were the only cells among those studied in which there was increased somatostatin inhibition of forskolin stimulated cyclase activity under elevated hydrostatic pressure (Fig 5B).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Percentage of stimulated adenylyl cyclase activity in different ocular cells exposed to elevated hydrostatic pressure. (A) Stimulation of adenylyl cyclase activity by forskolin (FSK). (B) Inhibition of forskolin stimulation by somatostatin (S). NPE = non-pigmented ciliary epithelium; PE = pigmented ciliary epithelium; TM = trabecular meshwork cells; A = lamina cribrosa astrocytes; R = retina cells.
Discussion
Our studies demonstrate that different cell lines derived from ocular tissues exhibit morphological and cellular activity responses following exposure to acute and sustained elevation of hydrostatic pressure to 50 mm Hg for up to 6 hours. Both phase contrast microscopy in living cells and cytoskeleton examination after phalloidin staining demonstrated that exposure to elevated hydrostatic pressure caused our ocular cells to appear more rounded in shape accompanied by changes in their actin cytoskeleton. In most cells in culture, the actin cytoskeleton is organised as microfilaments, which represent polymerised F-actin. The F-actin microfilament network can be observed at the microscopic level as stress fibres. In accordance with our observations, exposure to physiological or supraphysiological levels of hydrostatic pressure has been reported to cause reorganisation of stress fibres and profound changes in cell shape in a variety of cell types.18-24
Although exposure to elevated hydrostatic pressure did not affect cell viability, cellular activity of the intracellular enzyme adenylyl cyclase was notably affected. The basal adenylyl cyclase activity of each cell line significantly increased (∼50–80%) when incubated under elevated hydrostatic pressure. Maximum adenylyl cyclase activity in the presence of forskolin was calculated as the ratio of stimulated cyclase activity to basal activity. Therefore, decreased forskolin stimulation of adenylyl cyclase activity in all cell lines studied in the presence of elevated hydrostatic pressure is probably related to their increased basal adenylyl cyclase activity.
We observed a distinct difference in the astrocyte response to somatostatin treatment in comparison with the other cell lines studied. Specifically, astrocytes demonstrated the least amount of somatostatin inhibition of forskolin stimulated cyclase activity compared with other cell lines studied. Adenylyl cyclase activity is well known to be modulated by the activity of the stimulatory and inhibitory guanine nucleotide binding proteins, Gs and Gi.25 Since it is often difficult to assess accurately small changes in Gi activity by measuring the inhibition of basal adenylyl cyclase activity with drugs such as somatostatin, somatostatin inhibition of forskolin stimulated cyclase is generally used to amplify the Gi signal.16Our findings suggest that lamina cribrosa astrocytes behave differently with respect to the efficacy of receptor coupled Giactivity under either normal or elevated hydrostatic pressure conditions. This observation is in agreement with previous studies that somatostatin inhibition of stimulated cAMP accumulation provoked by other effectors such as forskolin and isoproterenol is weak in astrocytes.26 Although the difference between the response of astrocytes to somatostatin treatment in the presence or absence of elevated hydrostatic pressure did not reach statistically significant levels, astrocytes were the only cells among those studied that demonstrated increased somatostatin inhibition of forskolin stimulated cyclase activity under elevated hydrostatic pressure. Therefore, it appears that astrocytes may behave differently with respect to the regulation of the Gs and Gi “tone” that governs their adenylyl cyclase activity.
The elevated hydrostatic pressure model used in our experiments in which 50 mm Hg pressure was applied for up to 6 hours represents acute pressure elevation with a sustained component. Previous studies demonstrate that functional cell properties may be altered after brief exposure to elevated hydrostatic pressure. For example, cultured astrocytes open stretch activated channels half maximally after extremely brief (that is, seconds) exposure to hydrostatic pressure of 45 mm Hg.27 Furthermore, hydrostatic pressure (60–120 mm Hg) applied to astrocytoma cells for 30–60 minutes induces cell proliferation and DNA synthesis by activation of tyrosine kinase.28 Our data suggest that elevated hydrostatic pressure application for several hours can alter intracellular enzyme activity as well as perturb the actin cytoskeleton in ocular cells in vitro. We therefore hypothesise that functional cellular changes in response to brief elevations in external pressure, combined with the recovery that occurs following cessation of elevated pressure in vitro may result in significant changes in the homeostatic functions of several intraocular tissues.
The precise mechanisms of how the cells sense their environment and transduce a physical stimulus such as elevated hydrostatic pressure into a biochemical event, which may eventually lead to morphological and functional cellular changes are unclear. Mechanotransduction is thought to involve a coordinated system of molecules including extracellular matrix receptors, cytoskeletal proteins, and membrane channels.29-31 For example, hydrostatic pressure influences the action potential of the plasma membrane and produces depolarisation or hyperpolarisation, depending on the frequency and amplitude of the applied pressure.30 The plasma membranes can subsequently respond by increasing synthesis of cAMP,31 the cyclic nucleotide that is formed from ATP in reactions catalysed by adenylyl cyclase. Cyclic nucleotides such as cAMP play an important part in the normal metabolism and function of several ocular tissues including ciliary epithelium, trabecular meshwork, retina, and optic nerve.16 32-36 Therefore, increased basal adenylyl cyclase activity in ocular cells exposed to elevated hydrostatic pressure in vitro may reflect a characteristic cell adaptation of ocular cells to elevated intraocular pressure in order to maintain cell survival and function.
The most likely targets for cellular recognition of changes in hydrostatic pressure are the sites where the cell membrane interacts with underlying substrate and with neighbouring cells. The attachment sites of cells to the extracellular matrix and to the neighbouring cells through surface adhesion molecules in turn are connected to the cytoskeleton. One mechanism by which elevated hydrostatic pressure affects the cytoskeleton is thought to be related to regulatory mechanisms such as phosphorylation pathways that control cytoskeletal organisation triggered by activation of adenylyl cyclase in the cell membrane. For example, increased intracellular levels of cAMP and its effector, protein kinase A, can facilitate a reorganisation of the cytoskeleton, yielding rounded cells with short processes37—in accordance with our findings. The new cytoskeletal organisation observed in the cells exposed to elevated hydrostatic pressure may be a protective effort of these cells since it counteracts the initial membrane changes and potentially strengthens cytoskeletal integrity.31 38
In addition, our findings may have broad relevance to the physiological function of the secretory epithelium as they pertain to patients with glaucoma. Recently, it has been proposed that the rate limiting step in active secretion of aqueous humour by the ciliary epithelium is Cl ion transport via a 4,4′-diisothiocyanostilbene-2,2′disulphonate sensitive, cAMP dependent chloride channel.39 40 If elevated hydrostatic pressure, or more importantly, elevated intraocular pressure as occurs in patients with glaucoma, alters adenylyl cyclase activity in the ciliary epithelium as suggested by our findings, it seems quite feasible that cAMP dependent chloride transport, hence aqueous secretion, may be a dynamic process that is directly affected by the pressure level within the eye. This consideration has not yet been addressed in transport studies of the ciliary epithelium, which are typically performed at atmospheric pressure.
In summary, our observations provide evidence in five different cell lines derived from ocular tissues that exposure to sustained levels of hydrostatic pressure increases the level of adenylyl cyclase activity, in addition to reorganisation of the cell cytoskeleton and changes in cell shape. These in vitro observations suggest that key intracellular signalling cascades in ocular cells may be similarly affected by elevated intraocular pressure in vivo. Further studies seem warranted in order to assess other constitutive and altered functional responses of ocular cells to elevated pressure.
Acknowledgments
This study was supported in part by EY06810, EY12314, EY06416, and EY02687 from the National Institutes of Health, Bethesda, MD, the Glaucoma Research Foundation, San Francisco, CA, USA, and an unrestricted grant to Washington University School of Medicine, Department of Ophthalmology and Visual Sciences from Research to Prevent Blindness Inc, New York, USA.