Article Text

Abstract
Background: Because vascular endothelial growth factor (VEGF) signalling is deregulated in diabetic retinopathy, the potential therapeutic effects of VEGF inhibitors such as the human VEGF-specific antibody ranibizumab are currently being tested. A study was undertaken to determine whether VEGF-stimulated processes in retinal endothelial cells are reversed by ranibizumab.
Methods: The influence of VEGF121 and VEGF165 on the proliferation and migration of immortalised bovine retinal endothelial cells (iBREC) was studied in the presence and absence of ranibizumab. In addition, the protein composition of tight junctions in the presence of VEGF and its inhibitor in iBREC was investigated.
Results: While both isoforms stimulated proliferation of iBREC, only VEGF165 influenced cell migration. The addition of ranibizumab counteracted this stimulation without inhibition of the basal levels of migration and proliferation. Plasma membrane staining of the tight junction proteins occludin and claudin-1 disappeared in the presence of VEGF165; there was no effect on claudin-5 and ZO-1 was only weakly affected. The addition of ranibizumab restored plasma membrane localisation of occludin and claudin-1. For claudin-1, the variation in total protein expression corresponded with the observed effects of VEGF165 and ranibizumab.
Conclusion: Ranibizumab reverses proliferation and cell migration stimulated by VEGF and delocalisation of tight junction proteins induced by VEGF165 in iBREC.
Statistics from Altmetric.com
Diabetic retinopathy (DR) is one of the major complications of diabetes mellitus and the main reason for blindness in industrialised countries. It is generally accepted that the pathogenesis of DR is driven by deregulated expression of growth factors. Of these, vascular endothelial growth factor (VEGF) is considered most important as its levels are substantially raised in the vitreous fluid and in the retinal vasculature of patients with proliferative DR.1–4 VEGF not only influences the proliferation or migration of macrovascular and microvascular endothelial cells (EC) resulting in neoangiogenesis, but also increases the permeability of existing vascular surfaces leading to the breakdown of the blood-retina barrier as shown in vitro and in vivo.1 5–10 VEGF121 and VEGF165, the most intensively studied of the several isoforms of VEGF, bind to VEGF receptors 1 and 2 as well as to neuropilin 1, but the effects of VEGF on cellular proliferation, migration and permeability are mainly mediated through VEGF receptor 2.11 12 The increase in the permeability of retinal EC caused by VEGF165 is, at least in part, due to intracellular redistribution of occludin.13 14 This protein is a component of protein complexes which form so-called tight junctions of EC. These are involved in the control of the paracellular signalling pathway of these cells.15 16 In view of the strong evidence supporting a crucial role for VEGF, it seems reasonable to test drugs which interfere with VEGF-mediated signalling—such as the VEGF aptamer pegaptanib which interacts specifically with VEGF165 or the humanised Fab fragment ranibizumab which binds to all VEGF isoforms—as potential therapeutic options in the treatment of macular oedema.17 18
Neoangiogenesis in the retina is driven by proliferation and migration of retinal microvascular EC (REC) which are also the main constituents of the blood-retina barrier. Despite the crucial role of REC in the pathogenesis of DR, the effect of VEGF inhibitors on this cell type has not been studied. We have therefore investigated how migration, proliferation and tight junction composition in bovine retinal endothelial cells are affected by VEGF121 and VEGF165, two isoforms with an overlapping but not necessarily identical spectrum of physiological activities. In addition, we studied the potential inhibition of the effects of VEGF by ranibizumab, which was of particular interest in view of the emerging therapeutic relevance of VEGF inhibitors.
MATERIALS AND METHODS
Antibodies, reagents and media
Rabbit polyclonal antibodies binding to human claudin-1 (JAY.8), claudin-5 (Z43.JK), occludin (Z-T22) and ZO-1 (Z-R1) and with AlexaFluor 594 conjugated detection antibodies (F(ab′)2) were obtained from Invitrogen (Karlsruhe, Germany). Horseradish peroxidase-conjugated anti-rabbit secondary antibodies were included in the Opti4CN Detection Kit from Biorad (Munich, Germany). The Complete EDTA-free protease inhibitor cocktail was obtained from Roche Diagnostics (Mannheim, Germany) and the Phosphatase Inhibitor Cocktail 2 from Sigma-Aldrich (Deisenhofen, Germany). Recombinant human growth factors rhVEGF165 (SF21-expressed) and rhVEGF121 (SF-expressed) were from R&D Systems (Wiesbaden, Germany) and Calbiochem (Merck Darmstadt, Germany), respectively. Complete microvascular endothelial growth medium (ECGM; Promocell, Heidelberg, Germany) was supplemented with premixed additives resulting in final concentrations of 0.4% ECGS/H, 5% fetal calf serum, 10 ng/ml epidermal growth factor, 1 g/l glucose, 10 mM MgCl2 and 2.8 μM hydrocortisone. Basal medium (EBGM, Promocell) was supplemented only with either 103 nM (medium EBGM-lHC) or 2.8 μM hydrocortisone (EBGM-hHC). Ranibizumab (Lucentis) was a generous gift from Novartis Pharma GmbH (Nuremberg, Germany).
Cell cultivation
The propagation of immortalised microvascular endothelial cells from bovine retina (iBREC) established by transfection of primary retinal endothelial cells with plasmid pCI-neo hTERT encoding the catalytic subunit of human telomerase reverse transcriptase has been described previously.10 19
WST-1 cell proliferation assay
Cells (5×103 iBREC) were allowed to attach to the fibronectin-coated (50 μg/ml; BD Biosciences, Heidelberg, Germany) bottom of a cavity of a 96-well plate in ECGM overnight. After cultivation for 24 h in EBGM-hHC, VEGF121 or VEGF165 (1–100 ng/ml) and ranibizumab (1–100 μg/ml) as an optional supplement were added to the cells for 24 h before enzymatic conversion of WST-1 (Roche), indicative of proliferating cells, was measured according to the manufacturer’s protocol in eight replicate wells.
Cell migration assay
Transmembrane cell migration assays were performed in a modified Boyden chamber consisting of 12-well cell culture plates and inserts with a porous membrane (pore size 8.0 μm, diameter 1 cm; BD Biosciences). The lower compartment of the 12-well plate was filled with EBGM-hHC containing 5 μg/ml fibronectin, 1–100 ng/ml VEGF121 or VEGF165 and/or 5–50 μg/ml ranibizumab. Inserted membranes were initially incubated for 1 h at 37°C before 400 μl EBGM-hHC were added to the upper compartment. A suspension of 105 iBREC in 100 μl of this medium was then placed on the membrane and migration of cells through the pores of the membrane was assessed after 20 h at 37°C as described elsewhere.20
Immunofluorescence staining
iBREC were allowed to adhere to two-chamber slides (BD Biosciences or Nunc, Wiesbaden, Germany) coated with fibronectin in ECGM with 103 nM hydrocortisone overnight before they were cultivated in EBGM-lHC supplemented with 0.4% ECGS/H for 2 days. 10–100 ng/ml VEGF121 or VEGF165 were then added for 2 days and, subsequently, 100 μg/ml ranibizumab for an additional 24 h. Cells were fixed in methanol/acetone at −20°C and antigens were detected as described elsewhere.20 Primary antibodies against claudin-1 (5 μg/ml), claudin-5 and ZO-1 (both 2.5 μg/ml) and occludin (20 μg/ml) were diluted in 1% donkey serum in PBSd.
Western blot analyses
iBREC were subsequently treated with VEGF165 and ranibizumab as described above, with the exception that 0.1% FCS was added to the medium. To prepare whole cell extracts, 2×106 cells were detached by scraping, resuspended in 300 μl cold lysis buffer (40 mM TrisCl, 150 mM NaCl, 1% Brij 97, pH 7.4, supplemented with EDTA-free protease inhibitor and phosphatase inhibitor cocktails) and incubated on ice for 1 h. After centrifugation (10000×g, 4°C) for 15 min, Western blot analyses of tight junction proteins in the supernatant were performed as described elsewhere.10
Statistical analyses
All experiments in which data were generated from multiple replicates were repeated at least three times. The Mann-Whitney U test was used to analyse experimental data and resulting p values <0.05 were considered indicative of significant differences. The results are presented in conventional box-whisker diagrams with the middle line representing the mean and the borders of the boxes the 75% and 25% percentiles.
RESULTS
iBREC were chosen as a model to investigate the effect of ranibizumab on VEGF121- and VEGF165-stimulated processes because this homogenous cell line is free of contaminating cells and behaves like parental ECs of the bovine retina.10 In all experiments, concentrations of VEGF were in a range similar to those found in the vitreous fluid of individuals with proliferative DR, and the concentrations of ranizibumab reflected therapeutically achievable values.
Effect of ranibizumab on VEGF-induced iBREC proliferation and migration
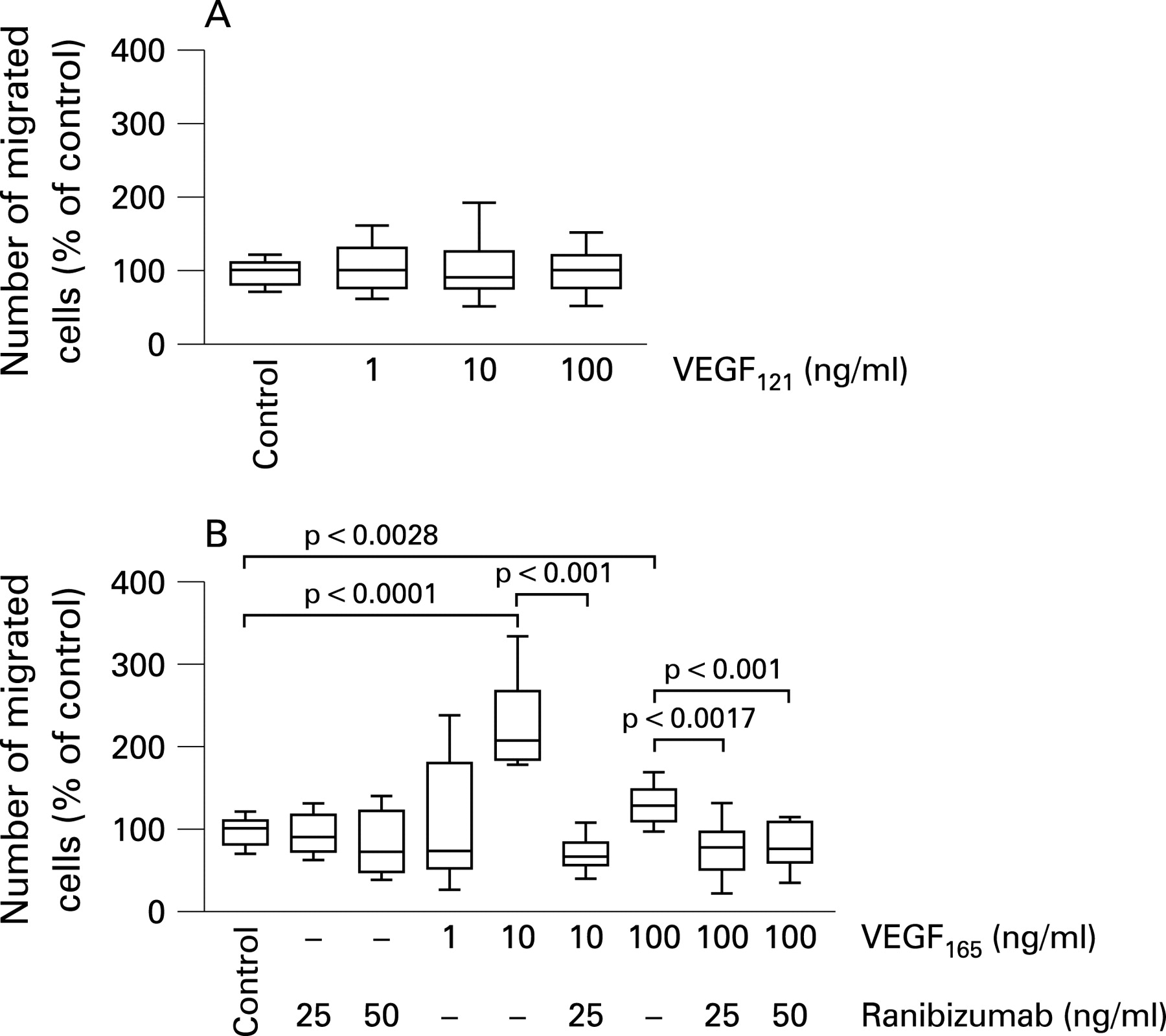
Serum-starved iBREC were exposed to VEGF121 or VEGF165 and ranibizumab at various concentrations for 24 h and conversion of WST-1 was measured as a means of cell proliferation (figs 1 and 2). The proliferation of iBREC, which had been significantly stimulated by VEGF121 at 1–100 ng/ml, was reduced to normal levels by treatment with 100 μg/ml ranibizumab; there was no effect on the proliferation of unstimulated cells (fig 1). Similar results were obtained with VEGF165 (fig 2). Concentrations of ranibizumab below 100 μg/ml were not sufficient to reverse VEGF121- or VEGF165-stimulated iBREC proliferation.


The migration of iBREC towards fibronectin in the presence of the different VEGF isoforms in the presence or absence of ranibizumab was studied in a modified Boyden chamber assay. As shown in fig 3, only VEGF165 stimulated cell migration in a concentration-dependent manner, whereas VEGF121 had no effect. Maximum stimulation was observed at a concentration of 10 ng/ml (2.5-fold, p<0.001), and the addition of 25 μg/ml or 50 μg/ml ranibizumab was sufficient to reverse the increased iBREC migration induced with 10 ng/ml or 100 ng/ml VEGF165, respectively.

Effect of ranibizumab on VEGF165-induced effects on tight junction proteins
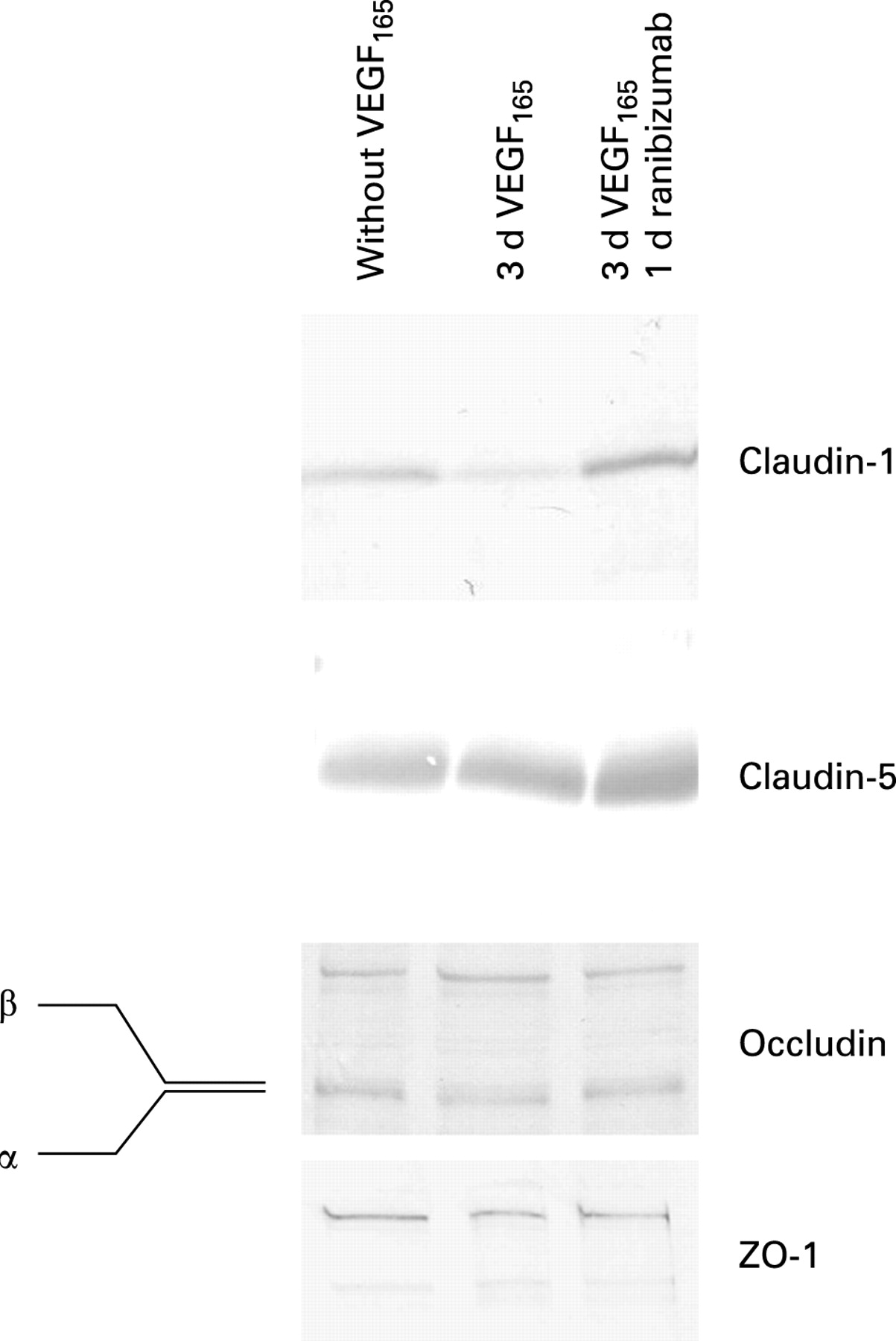
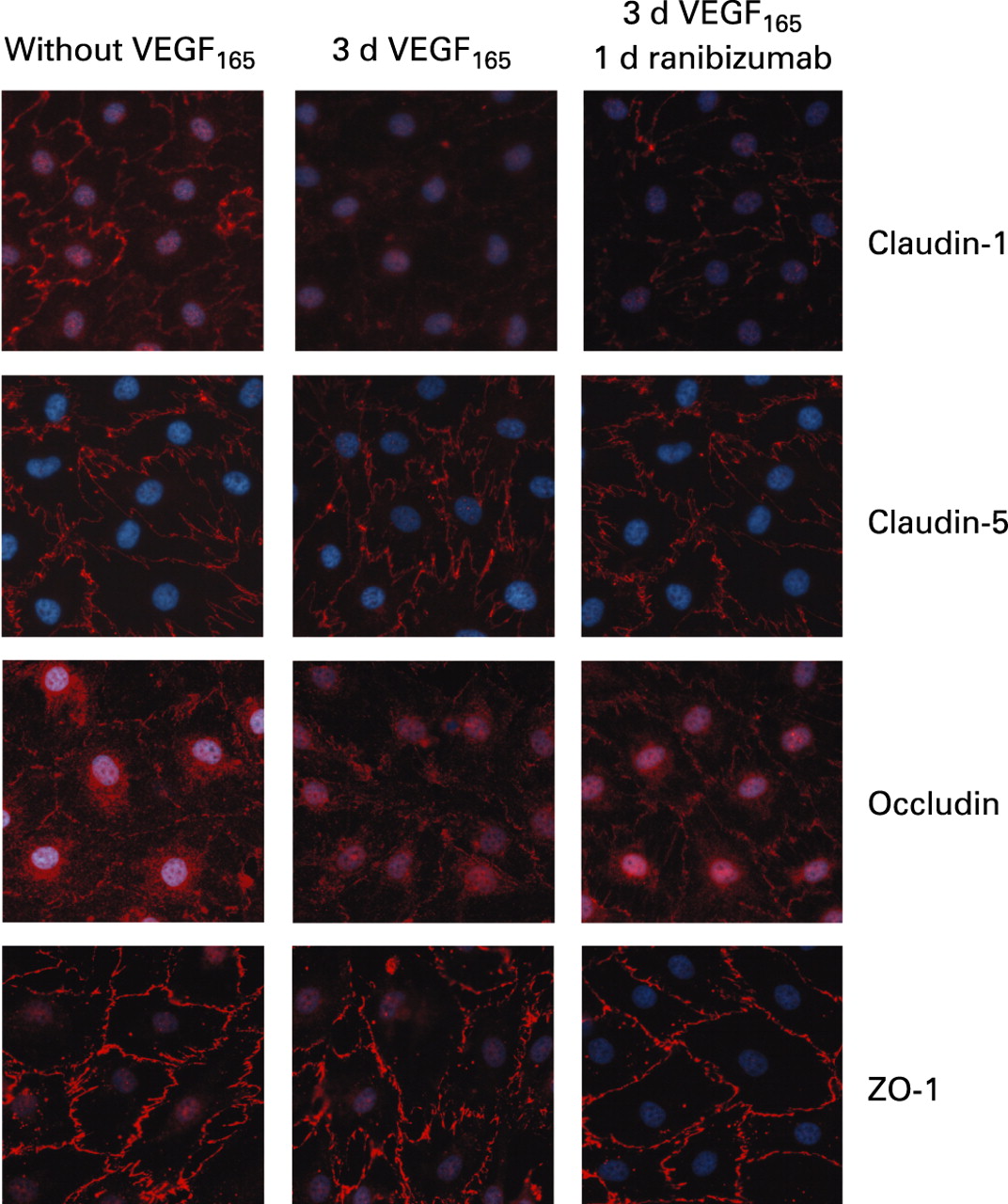
The tight junction proteins occludin, claudin-1, claudin-5 and ZO-1 were expressed by iBREC as shown by immunofluorescence staining and Western blot analysis (figs 4 and 5), whereas claudins 2 and 4—which are specific for epithelial cells—could not be detected in this cell type. In untreated confluent cultures, all these proteins were stained with variable intensities in the plasma membrane. The expression of occludin in the plasma membrane was strongly dependent on the concentration of hydrocortisone in the medium and was strongest at 100 nM, whereas no staining was evident after cultivation in medium with high (3 μM) concentrations or without hydrocortisone. In order to investigate the long-term effect of VEGF, iBREC were incubated with each isoform for up to 3 days. VEGF121 (10 and 100 ng/ml) or low concentrations of VEGF165 (10 ng/ml) did not markedly influence localisation of tight junction proteins. In contrast, after incubating the cells with 100 ng/ml VEGF165 for 3 days, plasma membrane localisations of claudin-1 and occludin were substantially weaker or even absent, which was often observed together with intensified intracellular staining of occludin (fig 4). The expression of ZO-1 at the plasma membrane was only slightly reduced after VEGF treatment. Plasma membrane localisation of claudin-5, which is specifically expressed by microvascular EC, was not influenced by VEGF. Incubation of VEGF165-treated iBREC in the presence of 100 μg/ml ranibizumab re-established plasma membrane localisation of all tight junction proteins tested within 24 h. Loss of plasma membrane staining of claudin-1 and occludin was seen as early as 30 min after the addition of VEGF165. While claudin-1 staining remained absent during extended cultivation, partial relocalisation of occludin occurred after treatment for 3 h.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Western blot analyses showed no change in the expression and modification of the proteins occludin, claudin-5 and ZO-1 in whole cell extracts from iBREC treated with VEGF165 for 2 days and subsequently with ranibizumab for 1 day (fig 5). However, the amounts of claudin-1 in these protein extracts were reduced after treatment with VEGF165 and increased again during subsequent cultivation with ranibizumab (fig 5). This variation in claudin-1 expression correlated with its delocalisation from the plasma membrane and reappearance after the addition of ranibizumab (figs 4 and 5). In agreement with results published by others, the higher phosphorylated β-form of occludin appeared to be slightly more prominent after treating iBREC with 100 ng/ml VEGF165 for 15–30 min (data not shown).13 14 However, after incubation for 3 h, both forms were again present in a ratio similar to that of untreateded cells (fig 5), indicating that phosphorylation of occludin was not stably affected by prolonged treatment with VEGF165.
DISCUSSION
Since deregulation of VEGF is considered one of the most important factors in the development of DR, influencing several important processes in retinal EC, VEGF inhibitors were evaluated as a possible therapeutic option. Although ranibizumab and the complete human anti-VEGF antibody bevacizumab have been shown to inhibit VEGF-induced proliferation of non-ocular human umbilical vein EC (HUVEC) and choroidal EC, similar studies have not been performed with retinal microvascular EC despite their physiological and clinical importance.21–24 We therefore studied the effect of VEGF and ranibizumab on cell migration, proliferation and composition of tight junctions in bovine retinal EC.
Cells of this type express VEGF receptors 1 and 2, and expression of neuropilin 1 can be induced by exposure to VEGF. Although the cells used in this study were of bovine origin, they responded to the human growth factors in a similar manner to that described for human retinal EC. In addition, immortalisation of bovine retinal EC by the introduction of hTERT did not result in any changes in the morphology, expression of cell type-specific proteins or induction of cellular processes by growth factors compared with parental primary cells.10 19 20 Because these cells do not contain any contaminating cells of other types, it can be expected that the influence of growth factors can be studied even more accurately and in greater detail in this model.
VEGF levels in vitreous fluid are raised in DR, even at early stages of the disease, and concentrations between 1 and 10 ng/ml have been reported.1–4 Maximum stimulation of iBREC proliferation and migration by VEGF isoforms was observed in a similar range of VEGF concentrations. Ranibizumab was able to inhibit these VEGF-induced processes to restore normal values without markedly influencing the basal levels at an antibody concentration of 100 μg/ml, which reflects the therapeutically achievable value calculated for an injection of 0.5 mg per eye, although local concentrations might be higher shortly after injection.25 Ranibizumab has been proposed to have antiproliferative effects on choroidal EC or ARPE19 cells which, however, might be due to traces of VEGF present in the medium originating from serum or endogenous production.22 Only VEGF165 stimulated migration of iBREC, although both VEGF121 and VEGF165 can induce migration of HUVEC, indicating the substantially different behaviour between macrovascular and microvascular EC.12 This might be due to different expression of the various VEGF receptors, leading to different activation of linked signalling pathways.
The permeabiltiy of retinal EC is increased after treatment with VEGF165, which probably leads to the breakdown of the blood-retina barrier observed in patients with DR from an early stage of the disease.5–8 13 In vitro studies have shown that bevacizumab inhibits VEGF-induced permeability of choroidal EC, providing a rationale for its therapeutic use.24 The pathway of paracellular signalling of EC is regulated at tight junctions, which consist of transmembrane proteins such as occludin and claudins as well as membrane-associated proteins such as ZO-1.15 16 The composition of these proteins determines the rate of transition of molecules through tight junctions, and their reorganisation—including delocalisation of components—is the most likely cause of the DR-related changes in permeability.13 14 In accordance with this assumption and the crucial role of VEGF, the amounts of claudin-1 and occludin localised in the plasma membrane were strongly reduced in iBREC after treatment with VEGF165 for 2–3 days. Ranibizumab re-established the plasma membrane localisation of claudin-1 and occludin in VEGF165-treated cells within 24 h, suggesting that this therapeutic agent can restore the functional integrity of tight junctions and subsequently readjust the increased permeability of iBREC. It is therefore assumed that ranibizumab might act in a similar way in vivo on the endothelium in the diabetic retina, with a therapeutic effect on macular oedema by directly counteracting its molecular causes.26
It has been shown that phosphorylation of occludin, by which its cellular localisation is most probably regulated, occurs shortly after treatment with VEGF165.13 14 27 28 In addition, the results of studies in an animal model indicate a slight increase in phosphorylated occludin after induction of DR.14 However, our results suggest that some difference in the degree of phosphorylation of occludin might influence its cellular localisation only immediately after stimulation, but this is probably not responsible for the altered localisation of this protein as a consequence of long-term treatment with VEGF165 in vitro. Highly phosphorylated occludin in epithelial MDCK I cells seems to be resistant to solubilisation with 1% NP-40.28 29 In iBREC, variation in the conditions for solubilisation did not yield different fractions of phosphorylated forms of occludin after treatment with VEGF165 for 3 days. However, part of the highly phosphorylated occludin was found in the Brij 97-insoluble fraction, but differences between VEGF165-treated and untreated iBREC were not observed (data not shown). Interestingly, in addition to the effects on occludin, we found that the expression of claudin-1 in iBREC was diminished by VEGF165, which was probably the reason for its disappearance from the plasma membrane, an effect that could be reversed by treatment with ranibizumab. It can be concluded that, although both components of tight junctions are at least in part co-localised in iBREC (data not shown) and the observed effects of VEGF165 and ranibizumab on these proteins occur synchronously, the underlying regulatory mechanisms are different for occludin and claudin-1.
This study, based on an in vitro model, confirmed that VEGF isoforms influence cellular processes which are likely to be responsible for the changes observed in the diabetic retina. The anti-VEGF antibody ranibizumab not only inhibited induction of these processes, but also reversed VEGF-induced delocalisation of tight junction proteins in iBREC, supporting the use of this drug in the treatment of DR. Ranibizumab potentially binds and neutralises all isoforms of VEGF, and all biological effects of VEGF including its role in the pathogenesis of DR should therefore be affected. Confirmation that ranibizumab reverses VEGF-induced changes in vitro is important because it not only confirms the dominant role of this growth factor in the progression of DR, but also shows that the binding characteristics and other properties of this antibody are sufficiently good to achieve a strong and pharmacologically relevant response. Ranibizumab might also prove to be a valuable tool for investigating the mechanism of action of the different VEGF isoforms, thereby fostering the development of small molecule drugs with similar activities.
Acknowledgments
The authors thank Anita Ruepp and Susanne Baur for expert technical assistance.
REFERENCES
Footnotes
Competing interests: This work was supported in part by an independent research grant from Novartis Pharma GmbH (Nuremberg, Germany).
Linked Articles
- At a glance