Article Text

Abstract
BACKGROUND/AIM It was previously reported that collagenous extracellular matrix (ECM) in human capsular opacification contained isoforms of transforming growth factor β (TGFβ). In the present study, the authors performed immunohistochemistry to examine whether ECM in human capsular opacification and in cultures of bovine lens epithelial cells (LECs) contained latent TGFβ binding protein-1 (LTBP-1), TGFβ1 latency associated peptide (β1-LAP), and fibrillin-1, a suspected ligand of LTBP-1 as well as a component of the extracellular microfibrillar apparatus. The aim of the study was to further clarify the mechanism of TGFβ1 deposition in ECM of capsular opacification.
METHODS Human capsular opacification specimens and uninjured lens capsules, as well as cultured bovine LECs, were processed for immunohistochemistry using antibodies against LTBP-1, β1-LAP, fibrillin-1, and collagen type I.
RESULTS LTBP-1, β1-LAP, and fibrillin-1 all were localised to the ECM in human capsular opacification. Uninjured lens epithelium stained for β1-LAP, but not for LTBP-1 and fibrillin-1. ECM deposited in confluent LEC cultures stained for LTBP-1, β1-LAP, and fibrillin-1, while cultures with only sparse cellularity were unstained for LTBP-1 or fibrillin-1.
CONCLUSIONS LECs upregulate LTBP-1 and fibrillin-1 during postoperative healing. LTBP-1, β1-LAP, and fibrillin-1 colocalised to the ECM in capsular opacification and in confluent LEC cultures. TGFβ1 is considered to deposit in ECM in the large latent form. ECM secreted by LEC may function as a scavenger or repository of TGFβ.
- posterior capsule opacification
- lens epithelial cell
- latent TGFβ binding protein-1
- fibrillin-1
Statistics from Altmetric.com
Capsular opacification is the most common complication of implantation of an intraocular lens (IOL). Lens epithelial cells (LECs) are responsible for this occurrence, depositing extracellular matrix (ECM) on the capsule.1-3 LECs may transform to a mesenchymal-like phenotype and produce ECM components including collagens, the major components of capsular opacification, in some conditions.4 5 LEC proliferation in association with ECM production is considered part of the wound healing process in the crystalline lens.
Transforming growth factor β (TGFβ) is pivotal in regulating cell behaviour during wound repair. Three isoforms (β1, β2, and β3) are known in mammals.6 7 Each isoform of TGFβ shows similar biological activities in vivo—that is, positive or negative regulation of cell growth, differentiation, and/or ECM production.8 Aqueous humour contains abundant TGFβ2.9 Moreover, healing LEC and ECM accumulation in capsular opacification reportedly contain TGFβ isoforms.10 11 Macrophages adhering to IOL are also immunoreactive for TGFβ isoforms.12 These isoforms are secreted as inactive complexes including small and large latent forms. Small latent TGFβ is composed of active TGFβ plus TGFβ latency associated peptide (β1-LAP); the large latent form consists of the small latent form of TGFβ plus latent TGFβ binding protein (LTBP). The ECM reportedly acts as a scavenger or repository for TGFβ in various tissues.13-15 Fibrillin-1, a component of the extracellular microfibrillar apparatus, reportedly can bind to LTBP-1.16 17 These two proteins share a number of features including multiple EGF-like repeats, the RGD sequence, and an eight cysteine repeat motif unique to this family of proteins.16 17 Immunolocalisation of LTBP-1 and fibrillin-1, in capsular opacification has not been fully examined, although TGFβ isoforms are located to LECs and/or ECM there.11 Fibrillin-1 is not a component of the normal lens, but may serve a connecter between LTBP-1 and ECM in capsular opacification similarly to that in ECM of other tissues. We therefore examined immunohistochemically localisation and protein expression patterns of LTBP-1, β1-LAP, and fibrillin-1 in human specimens with capsular opacification to further clarify the mechanism of TGFβ1 deposition in ECM of capsular opacification. Furthermore, we determined that cultured bovine LECs produce and deposit LTBP-1, β1-LAP, and fibrillin-1 in their ECM.
Materials and methods
CLINICAL SPECIMENS
Specimens obtained from 25 Japanese patients with capsular opacification were included in this study (Table 1). Three circular sections of the anterior capsule with LECs without opacification of the anterior subcapsule were obtained by circular capsulotomy during phacoextraction in patients with posterior subcapsular cataracts. Specimens from nine patients (cases 1–9) were obtained at the department of ophthalmology of Wakayama Medical University Hospital, while specimens from 16 patients (cases 10–25) were provided from the implant data system committee of the Japanese Society of Cataract and Refractive Surgery. The specimens were embedded in paraffin after being fixed in 10% formalin. Tissue sections 5 μm thick were cut, deparaffinised, and processed for immunohistochemistry as described below.
Summary of the cases and the results
CELL CULTURE
Lens capsules were obtained from five bovine eyes. The tissues were explanted for outgrowth of LECs in Eagle's medium supplemented with antibiotics, antimycotics, and 10% fetal calf serum.18 After one passage, the cells (5 × 104/ml/well) were seeded on chamber slides (Nunc, Naperville, IL, USA) and incubated for 3, 10, or 14 days. Three chambers were prepared for each culture condition. Cells then were dried, fixed in cold acetone, and processed for immunohistochemistry.
IMMUNOHISTOCHEMISTRY
Immunohistochemistry was performed as previously described.19 Deparaffinised sections and fixed cultured cells were treated with phosphate buffered saline (PBS) containing 3% skim milk and 5% calf serum to decrease non-specific binding of antibodies. Specimens then were allowed to react with the primary antibodies listed in Table 2 for 12 hours at 4°C. After washing in PBS, specimens were treated with peroxidase conjugated secondary antibodies for 4 hours at 4°C as previously reported.5 11 12 After a colour reaction with 3,3′-diaminobenzidine, specimens were embedded in balsam and observed by light microscopy. Negative controls were prepared by omission of the primary antibodies.
Primary antibodies used
Results
SPECIMENS OF UNINJURED LENS CAPSULE AND CAPSULAR OPACIFICATION
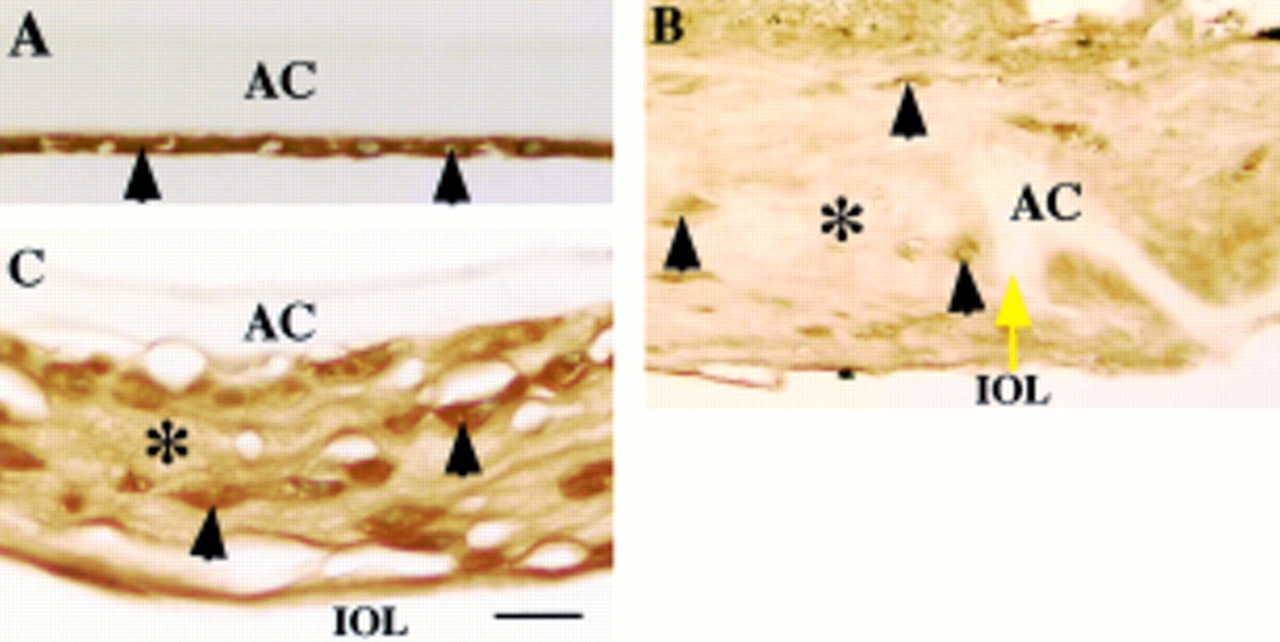
LTBP-1 was immunohistochemically detected in ECM accumulation in the lesion of human fibrous capsular opacification, but not in uninjured LECs (Fig 1). Immunoreactivity for β1-LAP was marked in the cytoplasm of both uninjured LECs and healing lens cells in the fibrotic capsular opacification (Fig 2). The ECM of fibrotic capsules also was also reactive for β1-LAP (Fig 2). The regenerated lenticular structure of Sommerring's ring and also healing lens cells were unstained for these two proteins (Fig 1D). LECs of uninjured lenses were negative for fibrillin-1, but LECs in healing capsular opacification were surrounded by fibrillin-1 deposition. Fibrillin-1 was immunohistochemically detected in ECM accumulation on fibrous capsular opacification (Fig 3). Similar results were obtained in fibrillin-1 immunohistochemistry with two primary antibodies of different sources. No immunoreactivity was observed in negative control staining (not illustrated). There is no difference of the expression patterns of these three proteins among specimens of uninjured lens cells of various ages in given specimens.
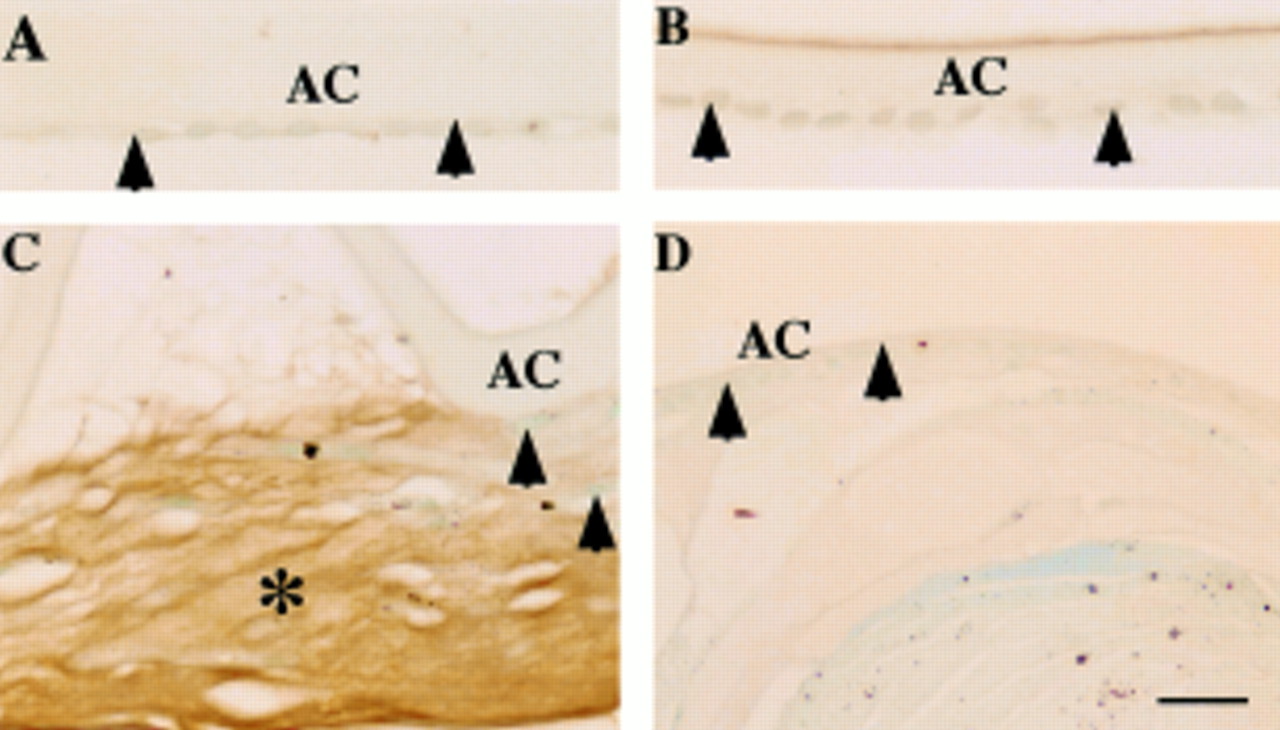
Localisation pattern of large transforming growth factor β binding protein-1 (LTBP-1) in human posterior capsule opacification. In (A) (case 1) and (B) (case 8), lens epithelium of an uninjured lens capsule (A, arrowheads) and a capsule 10 days after cataract surgery (B, arrowheads) lack LTBP-1. In (C) (case 17), extracellular matrix accumulation (asterisk) between the anterior lens capsule and the anterior surface of the intraocular lens shows immunoreactivity for LTBP-1. Lens cells are negative for this protein (arrowheads). In (D) (case 19), lens cells (arrowheads) and regenerated lenticular fibres of Sommerring's ring unstained for LTBP-1. AC = anterior capsule; IOL = intraocular lens. Indirect immunostaining; bar = 20 μm.

Localisation pattern of β1 latency associated peptide (β1-LAP) in human posterior capsule opacification. In (A) (case 1), lens epithelial cells (arrowheads) of an uninjured lens capsule exhibit immunoreactivity for β1-LAP. In (B) (case 25), extracellular matrix accumulation (asterisk) around the edge of the capsulotomy (yellow arrow) shows weak immunoreactivity for β1-LAP, while lens cells (arrowheads) exhibit marked immunoreactivity. In (C) (case 17), extracellular matrix accumulation (asterisk) between the intraocular lens optic and the anterior capsule as well as lens cells (arrowheads) show strong β1-LAP immunoreactivity. AC = anterior capsule; IOL = intraocular lens. Indirect immunostaining; bar = 20 μm.

Fibrillin-1 as a component of fibrous extracellular matrix in human posterior capsule opacification. In (A) (case 2), uninjured lens epithelium (arrowheads) lacks immunoreactivity for fibrillin-1. In (B) (case 18), extracellular matrix accumulation (asterisk) around the edge of anterior capsulotomy (yellow arrow) shows marked immunoreactivity for fibrillin-1. In (C) (case 17), weak immunoreactivity for fibrillin-1 (arrows) is detected surrounding proliferating lens epithelial cells, which lack immunoreactivity. AC = anterior capsule; IOL = intraocular lens. Indirect immunostaining; bar = 20 μm.
LEC CULTURE
In sparsely cellular preconfluent cultures at day 3, almost no immunoreactivity for LTBP-1 was detected in LECs (Fig 4A). In early confluence, a small amount of LTBP-1 was detected extracellularly in a very fine fibrillar pattern, while in mature confluent cultures at day 14 immunoreactivity for LTBP-1 appeared as an organised extracellular network throughout the culture (Fig 4B and C). Immunoreactivity for β1-LAP was detected in cells in sparsely cellular cultures and early confluent cultures (Fig 4D and E), although its immunoreactivity seemed weaker compared with that of in vivo LECs. Mature confluent cultures of LECs exhibited extracellular deposition of β1-LAP (Fig 4F). Sparsely cellular LEC cultures did not show fibrillin-1 protein expression (Fig4H), while LECs in early confluent cultures at day 10 were reactive for fibrillin-1 in a fine fibrillar pattern in the ECM (Fig 4I). The cells themselves remained unstained. Mature confluent LEC cultures showed a condensed fibrillar structure of fibrillin-1 in their ECM, remaining the cell cytoplasm negative (Fig 4J). Fibrillin-1 staining patterns did not completely overlap with immunoreactivity for LTBP-1 or β1-LAP. At all time points during culture, collagen type I was detected in the cytoplasm of LECs in a fine granular pattern, but not in the extracellular space (not illustrated). No specific immunoreactivity was observed in negative controls with anti-goat IgG or anti-rabbit IgG, but no primary antibody (Fig 4G and K).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Immunolocalisation of LTBP-1, β1-LAP, and fibrillin-1 in the extracellular matrix accumulation in bovine lens epithelial cell cultures. In sparse, preconfluent, cultures at day 3, almost no immunoreactivity for LTBP-1 was detected in LECs (A). In early confluence, a small amount of LTBP-1 was detected extracellularly in a very fine fibrillar pattern (B), while in mature confluent cultures at day 14, immunoreactivity for LTBP-1 appeared to form an organised network throughout the culture (C). Immunoreactivity for β1-LAP was detected in the cells of a sparsely cellular culture (D) and an early confluent culture (E). The mature confluent culture of LECs exhibited extracellular deposition of β1-LAP (F). In sparsely cellular cultures, LECs did not express fibrillin-1 protein (H). In early confluent cultures at day 10, fibrillin-1 was detected in a fine fibrillar pattern in fibroblast ECM; the cells themselves were unstained (I). Mature confluent LEC cultures showed a condensed fibrillin-1 structure (J). No specific immunoreactivity was observed in negative controls with anti-goat IgG (G) or anti-rabbit IgG but no primary antibody (K). LTBP-1 = large transforming growth factor β binding protein-1; β1-LAP = β1-latency associated peptide; LEC = lens epithelial cells. Indirect immunostaining; bar = 50 μm.
Discussion
In the present study, immunostaining patterns for LTBP-1, β1-LAP, and fibrillin-1 suggested that TGFβ1 and fibrillin-1 were colocalised to the ECM in human capsular opacification. Uninjured LECs were positive for β1-LAP but negative for LTBP-1 and fibrillin-1, indicating that in vivo LECs upregulate LTBP-1 and fibrillin-1 during healing after surgery. Fibrillin-1 is considered to trap TGFβ1 through binding to LTBP-1.We previously reported that myofibroblastic transformation of in vivo LECs during lens wound healing is associated with expression of various ECM macromolecules including collagens.5 LTBP-1 and fibrillin-1 are considered to be upregulated similarly to other ECM components expressed by LECs in association with myofibroblastic transformation, in that cuboidal LECs in a specimen 10 days postoperatively or in Sommerring's ring were negative for LTBP-1. Alternatively, ECM surrounding postoperative LECs may provide some signal(s) for expression of LTBP-1 (or for conversion from the small latent form to the large latent form of TGFβ1). Indeed, fibronectin reportedly regulates the epithelial mesenchymal conversion in cells including LECs, as shown by an α smooth muscle actin expression pattern.20 21 Similar mechanisms may work in upregulation of LTBP-1. Fibrillin-containing microfibrillar apparatus is considered to play an important part in ECM organisation during lens tissue repair, as occurs in other tissues.22 23 Localisation of LTBP-1 and β1-LAP to fibrous capsular opacification suggests that it functions in storing or scavenging latent TGFβ in this ECM compartment.
We further confirmed by cell culture that bovine LECs deposit LTBP-1 and β1-LAP as well as fibrillin-1 in the ECM. Deposition of LTBP-1, β1-LAP, and fibrillin-1 also has been demonstrated in other cell types in culture.13-17 In sparsely cellular LEC cultures, LTBP-1 and β1-LAP were not detected in the cell layer, but in confluent LEC culture they were observed in the ECM in a fibrillar network or a granular pattern. Cell signals induced by confluence apparently stimulate deposition of LTBP-1, β1-LAP, and fibrillin-1 into the extracellular space. In mature confluent cultures, fibrillin-1 containing structures did not completely overlap with immunoreactivity for LTBP-1 and β1-LAP, indicating that LTBP-1 and β1-LAP are deposited in areas where no fibrillin-1 is present in addition to being bound to fibrillin-1. Other depositing mechanism may also be present. In this stage of cultures, however, collagen type I, the major ECM component in fibrous capsular opacification, was detected only in cytoplasm, not in the ECM on the cells; thus, LTBP-1 and β1-LAP are not associated with type I collagen. Secretion of β1-LAP and fibrillin-1 by cultured LECs might not be similar to that in in vivo LECs; cytoplasmic immunoreactivity for both proteins were marked in the cytoplasm of healing LECs in vivo, but weak or not seen in cell culture. Further study is needed to clarify the regulation mechanism of secretion of these proteins.
LECs proliferate following cataract surgery even though they are exposed to TGFβs that have a growth inhibitory effect on them.24-29 ECM molecules may regulate (enhance) postoperative proliferation of LECs in vivo.30 We have reported that lens epithelial cells in capsular opacification express matrix metalloproteinases and their inhibitors.31Degradation activity of fibrillin-1 containing ECM may modulate LEC growth by regulating release of TGFβ from the matrix.
In conclusion, ECM accumulation surrounding LECs in vivo postoperatively and those in vitro deposits TGFβ1 presumably through binding LTBP-1 to fibrillin-1, and may modulate LEC behaviour not only by providing cell adhesion scaffolds but also by storage or scavenging of this growth factor.
Acknowledgments
Grant support: Supported in part by Wakayama Medical Award for Young Researchers, 1999.
The authors thank members of the IOL Implant Data System Committee of the Japanese Society of Cataract and Refractive Surgery for their cooperation.